


All Issues
Yield in almond is related more to the abundance of flowers than the relative number of flowers that set fruit
Publication Information
California Agriculture 71(2):68-74. https://doi.org/10.3733/ca.2016a0024
Published online November 16, 2016
NALT Keywords
Abstract
Almond tree yield is a function of the number of flowers on a tree and the percentage of flowers that set fruit. Almonds are borne on spurs (short proleptic shoots that can have both leaves and flowers). Almond tree spur dynamics research has documented that previous year spur leaf area is a predictive parameter for year-to-year spur survival, spur flowering and to a lesser extent spur fruiting, while previous year fruit bearing has a negative impact on subsequent year flowering. However, a question remained about whether yields are more dependent on flower numbers or relative fruit set of the flowers that are present. The aim of the present work was to compare the importance of flower abundance with that of relative fruit set in determining the productivity of a population of tagged spurs in almond trees over a 6-year period. Overall tree yield among years was more sensitive to total number of flowers on a tree rather than relative fruit set. These results emphasize the importance of maintaining large populations of healthy flowering spurs for sustained high production in almond orchards.
Full text
Almond is the most important tree nut crop in California in terms of acreage and production value (USDA NASS 2015). Grower attention has been focused on increasing orchard yields for decades (Kester and Griggs 1959) and research efforts have been aimed at improving almond orchard productivity through the optimization of all variables involved in nut production. Fundamentally, almond tree yields are the product of the number of kernels produced per tree and kernel weights. Of these two factors, the number of kernels is the most important (Reidel et al. 2001) since kernel weight is generally not a factor of paramount importance for growers (Spiegel-Roy and Weinbaum 1985).
The reproductive process in almond trees involves two years from flower bud induction to fruit set and fruit maturity. In this process the number of flowers borne and the number of flowers that set fruit determines the final kernel yield per tree. Fruit set in almond is strongly influenced by presence of pollinizer cultivars, insect pollinators and by climatic conditions affecting pollen viability, germination and pollinator activity (Corbet 1990; Dulberger et al. 1994; Eisikowitch et al. 1991; Gradziel and Weinbaum 1999; Hedhly et al. 2007; Kozlowski and Pallardy 2002; Ortega et al. 2007; Thorp 1996; Tombesi et al. 2010; Vasilakakis and Porlingis 1985; Weinbaum et al. 1984) in addition to general tree health.
In 1959, Kester and Griggs stated that “the question often arises as to whether or not the fruit set for specific almond orchards could be increased by using more bees and pollinizers to effect more complete cross pollination”. Mean relative fruit set (i.e., percentage of flowers that set a fruit) in almond has been reported to be about 30% but there is large year-to-year variability that can make it range as low as 5% (Socias i Company 1994) and as high as 40% (Kester and Griggs 1959). These relatively low fruit set percentages offer a potential margin for almond crop improvement. Accordingly, almond orchards are planned and managed to improve relative fruit set by planting pollinizer rows on either side of the main cultivar rows to increase availability of compatible pollen (Dag et al. 2000). The use of bees in almond orchards during flowering increases the likelihood of movement of pollen among trees (Artz et al. 2013; Brittain et al. 2013). Enhancing tree nutrition has also been reported to increase fruit set rates (Nyomora et al. 1997, 1999). In spite of these efforts, relative fruit set is still variable and little improved since the early data reported in 1959 by Kester and Griggs.

Results from spur dynamic studies indicate that maximizing healthy populations of productive spurs is key to optimizing yields.
Almond spur dynamics
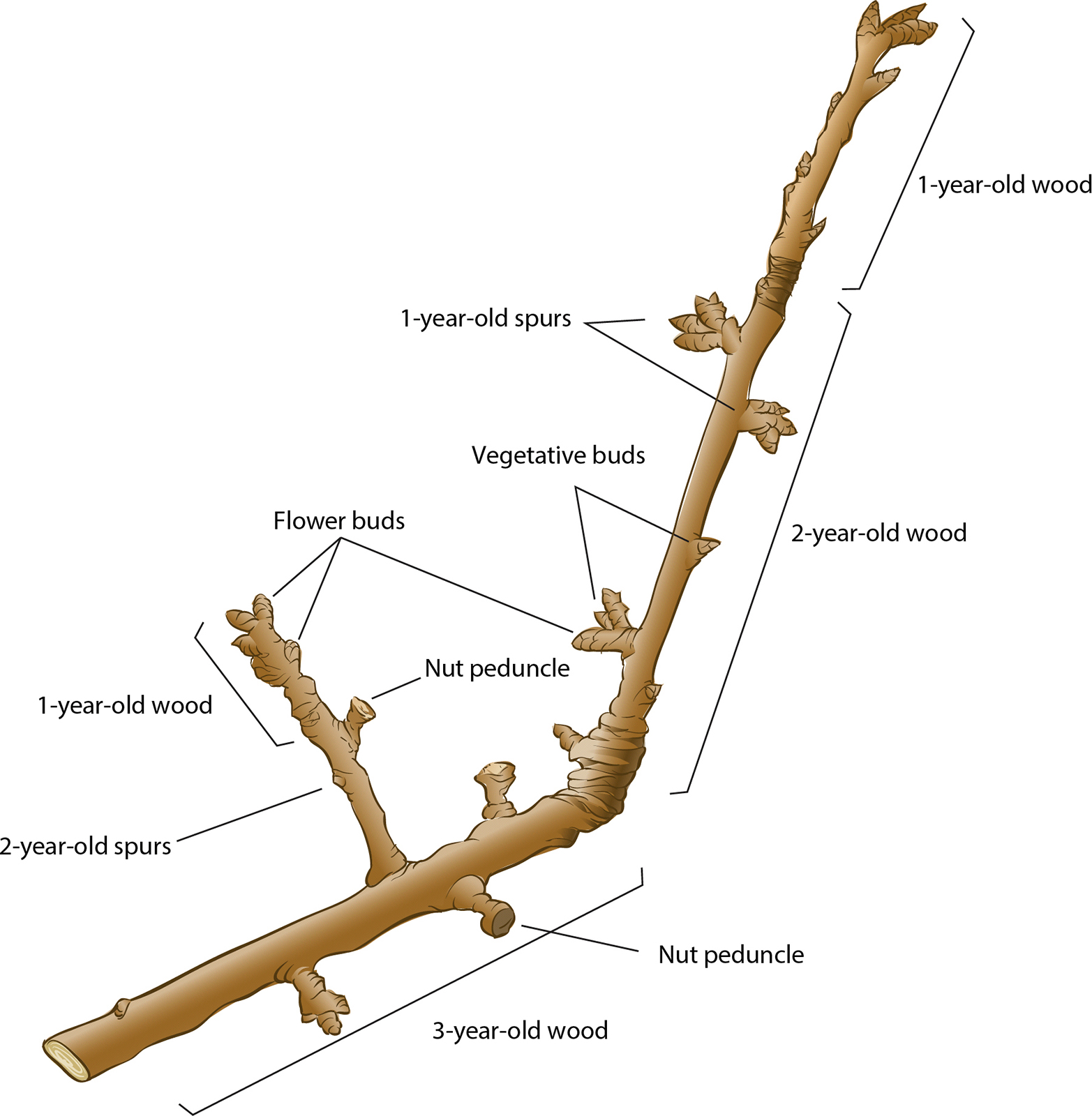
But, is fruit set the main limiting process for almond productivity? Another approach could be to increase the number of flowers per acre — but that approach demands more information on the eco-physiological basis that regulates flowering of almond spurs (short lateral shoots that are the main flowering and fruit bearing units in mature almond trees — see illustration). Individual spurs tend to alternate bear with only a small percentage of spurs flowering the year after bearing (Lampinen et al. 2011). The authors have observed tagged spurs in outer canopy–exposed positions to live at least 15 years.

An almond spur with a flower in full bloom. The number of flowers that set fruit determines the final kernel yield per tree.
To investigate this, an almond spur dynamics research project was initiated by Lampinen and colleagues in 2001. This study was designed to quantify the dynamics of spur renewal, fruitfulness and longevity and to determine how these dynamics are impacted by orchard management practices. Results from the study indicated that the number of flowers borne by individual spurs is a function of spur leaf area in the previous year and whether or not the spur bore a fruit in the previous year. Spurs that bore fruit in a given year rarely flowered or bore fruit in a subsequent year (Lampinen et al. 2011). Furthermore, spur mortality was much higher in spurs that had low previous year spur leaf area (PYSLA) because fruit bearing competes with leaf growth and decreases the amount of source organ available on bearing spurs (Lampinen et al. 2011; Tombesi et al. 2015). Although there was a strong tendency for individual spurs to not bear fruit in successive years, whole trees or orchards are not strongly alternate bearing because fewer than 20% of the spurs on a tree bear fruit in a given year (Tombesi et al. 2011).
In addition, the spur dynamics study documented that the key to ensure the largest flowering over an orchard's life is to have the largest number of spurs possible with the optimal leaf area for flowering. Proper irrigation during the previous year vegetative season and even after harvest can help to minimize spur death and has been reported to have a critical impact on subsequent bloom and fruit set (Esparza et al. 2001; Goldhamer and Smith 1995).
The almond spur dynamics study also provided information regarding the importance of PYSLA in determining subsequent spur flowering, fruit bearing and survival (Lampinen et al. 2011) as well as the fact that spur fruit bearing in turn, reduces spur leaf in the same year (Tombesi et al. 2015). Thus, spur flowering and fruiting in two sequential years is relatively rare (Tombesi et al. 2011). However, the total number of flowering spurs on a tree may be of limited significance if greater relative fruit set of the flowers can compensate for decreased flower numbers in the orchard. Thus, understanding the relative impact of flower number and relative fruit set on almond tree yield in commercial orchards is essential for guiding efforts to improve orchard productivity and help growers determine the most profitable practices for almond crop management.
Study of flowering and fruit set
To address this question we analyzed flowering and fruit set data recorded during the almond spur dynamics project. The study was conducted in a 145-acre orchard, planted in 1996, at 24 feet between and 21 feet within rows. The orchard planting consisted of rows of ‘Nonpareil’ (50%) alternating with pollinizer rows of ‘Monterey’ (25%), and ‘Wood Colony’ (25%). The orchard was located in Kern County on a sandy-loamy soil. Irrigation was carried out by microsprinklers and irrigation schedule was based on weekly measurement of midday stem water potential that was maintained between −0.7 and −1.2 MPa. Nitrogen was applied at 110 to 220 pounds per acre and leaf N content was between 1.95% and 2.45% over the period of the experiment. Bee hives were placed at a density of two to three hives per acre prior to bloom. During the experiment, weather conditions during the pollination period were not limiting for bee activity.

A vegetative almond spur, top, and fruitful spur, bottom, marked with aluminum tags. Spurs are short lateral shoots that are the main flowering and fruit bearing units in almond trees.
The orchard was divided into six equal-sized replicate blocks and 50 spurs were tagged in eight ‘Nonpareil’ trees within each of the six blocks. A total of 2,400 spurs were marked with aluminum tags in late March and early April 2001. Twelve spurs were selected on each of the northeast and northwest quadrants of individual trees and 13 spurs were selected on each of the southeast and southwest quadrants of the same trees. Tagged spurs were located at positions ranging from shaded (near the trunk) to exposed (on the periphery) portions of the canopy at a height of 3 to 12 feet. During the first 4 years of the study, lost tags or dead spurs were replaced with spurs in close proximity with similar light exposure to the original tagged spurs.
The dynamics of annual growth, flowering, fruitfulness and spur mortality were quantified annually. For more detail see Lampinen et al. (2011). The number of flowers produced on each tagged spur was counted in the spring of each year from 2002 through 2007. Multiple year records of PYSLA (from an adjacent, similar spur as described earlier), previous year bearing, number of flowers in the current year and number of fruit in the current year were used to assess spur behavior in relation to PYSLA in spurs that bore no fruits in the previous year. These analyses involved data from 6,980 spurs spread over the 6 years.
Kernel yield of the individual trees with tagged spurs and the kernel yield of the orchard containing those trees were also recorded for 6 years (2002–2007).
Statistical analyses were carried out using ANOVA (Sigmaplot 8.0, SPSS Inc., Chicago, Illinois) to test the significance at P < 0.01 of relationships between PYSLA and current year spur flower density (flowers per spur), current year spur fruit density (fruit per spur) and current year spur relative fruit set. The same test was also used to test the significance of the relationship between tree yield (expressed as kilograms of kernels per tree) and tree spur population relative fruit set (expressed as the relative fruit set recorded on the 50 spurs tagged on each tree) and spur flower density (expressed as the mean number of flowers per spur recorded on the 50 spurs tagged on each tree).

Effects on yield
The number of flowers differentiated (formed) during the previous year is the first component of yield in fruit trees (Werner et al. 1988). In almond spurs, flower formation was closely related to spur leaf area in the previous year (PYSLA) (fig. 1). Thus, if the leaf area of each spur on a tree were known, the number of flowers that a tree would bear in the following year could be estimated, and, if spur relative fruit set were constant, spur fruit bearing and yield of that tree could be predicted. However, although the relationship between spur fruit density and PYSLA was significant, it was weaker than the relationship between spur flowering and PYSLA (fig. 2). This was because fruit set was highly variable in almond across years. Relative fruit set varied from 19% to 36% (table 1).

Fig. 1. Relationship between current year spur flower density and previous year spur leaf area on tagged spurs from 2002 to 2007 (R2 = 0.76, P < 0.0001). Each point is the mean of 10 spurs ± SE.
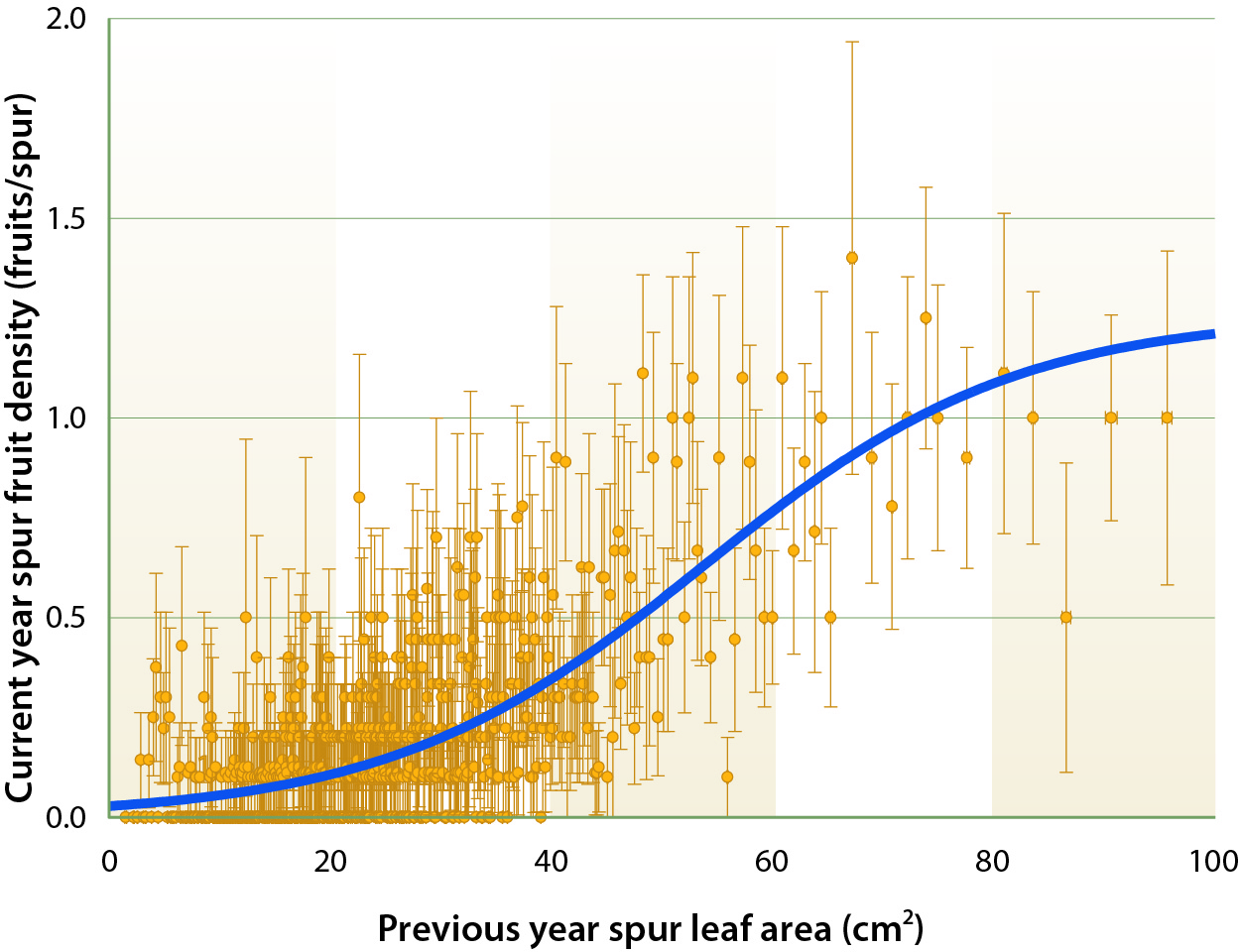
Fig. 2. Relationship between current year spur fruit density and previous year spur leaf area on tagged spurs from 2002 to 2007 (R2 = 0.59 P < 0.0001). Each point is the mean of 10 spurs ± SE. 1 cm2 = 0.001 ft2.

TABLE 1. Number of flowers, fruits and relative fruit set rate of the spur population evaluated from 2002 to 2007
These data apparently support the large effect of season, and particularly weather conditions, on the fruit set process. In almond, rainfall during the bloom period has been reported to affect pollinator activity (Eisikowitch et al. 1991; Vicens and Bosch 2000) and to wash pollen off stigmas (Dulberger et al. 1994; Ortega et al. 2007). Anther dehiscence (shedding of pollen by the anther) also can be affected by rain (Corbet 1990) and high relative humidity (Gradziel and Weinbaum 1999; Kozlowski and Pallardy 2002). Temperature affects pollen germination, pollen tube growth (Vasilakakis and Porlingis 1985; Weinbaum et al. 1984), ovule degeneration (Hedhly et al. 2007; Postweiler et al. 1985) and pollinator activity in the field (Corbet et al. 1993; Thorp 1996; Vicens and Bosch 2000). Wind can also affect pollinator activity (Thorp 1996; Vicens and Bosch 2000). On the basis of this information, some have hypothesized that yield fluctuations can be explained mainly by variations in climatic factors (Dorfman et al. 1988). Actually, large relative fruit set variability also occurred among individual trees (fig. 3). This fluctuation could be a result of “on-trees” and “off-trees” (i.e., bearing more or less nuts than their average production) occurring in the same orchard and season (Tombesi et al. 2011). On the other hand, fluctuations of relative fruit set of spur populations in different trees exposed to the same climatic conditions suggest that climatic conditions are not the major factor influencing tree spur population fruit set.
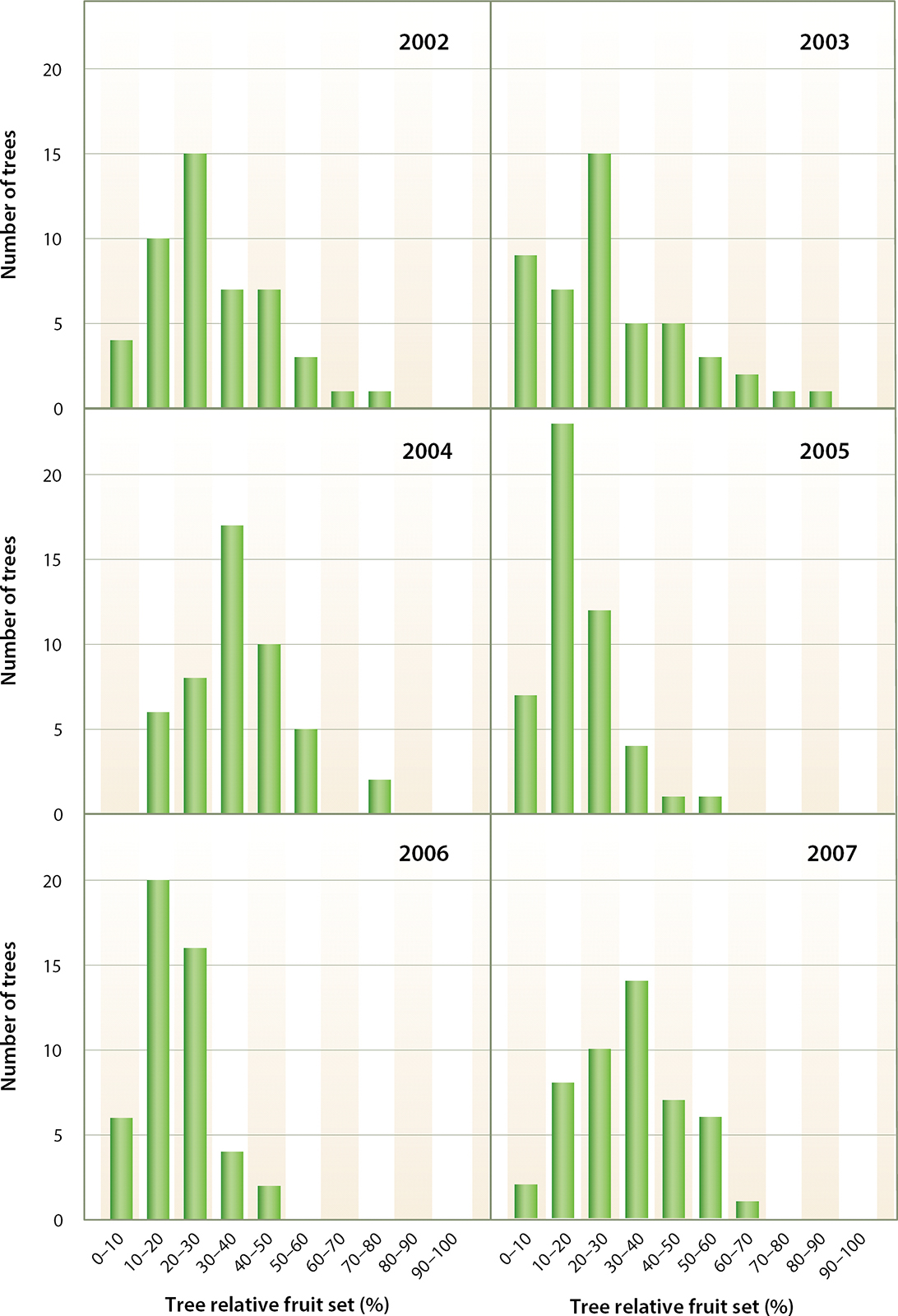
Fig. 3. Distributions of number of trees with different tree relative fruit sets from 2002 to 2007. Relative fruit set was recorded on 50 spurs per tree and was obtained as the number of fruit borne on all 50 spurs divided by the number of flowers borne on all the 50 spurs times 100.
In this experiment, at the spur level, there was no correlation between the PYSLA and relative fruit set in the current year (fig. 4). Thus, whereas previous year conditions are fundamental for flower formation on spurs (Lampinen et al. 2011), previous year leaf area did not appear to influence current year spur relative fruit set. Furthermore, spur fruiting was associated with reduced spur leaf area in the current season, suggesting that current year spur leaf area does not exert any influence on spur relative fruit set (Tombesi et al. 2015). In this experiment, the number of nuts borne by individual trees was significantly correlated with the number of nuts borne by the tagged spur populations in those trees (fig. 5). This suggests that our spur sample was relatively representative of the spur population of the trees. On a whole tree basis, tree yield was not correlated with mean relative fruit set measured on tree spur populations. Instead, tree yields appeared to be more closely correlated with flower density on the tagged spur population. Thus, while relative fruit set is obviously important, it was not the primary yield-limiting factor in this orchard situation, and increased relative fruit set when floral densities were low did not compensate for lower numbers of flowers (fig. 6).
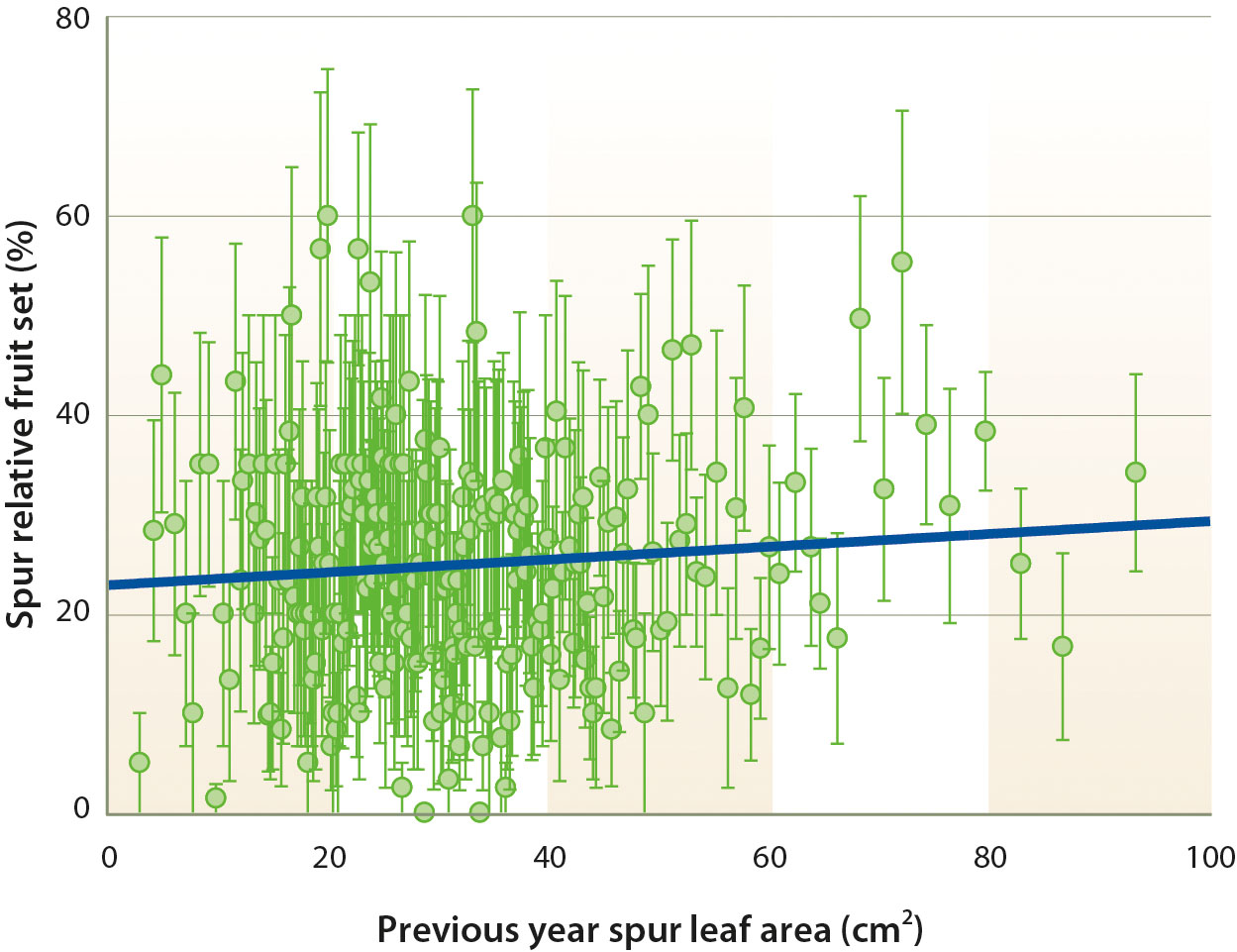
Fig. 4. Relationship between spur relative fruit set and previous year spur leaf area on tagged spurs from 2002 to 2007 (R2 = 0.007, P = 0.16). Each point is the mean of 10 spurs ± SE. 1 cm2 = 0.001 ft2.
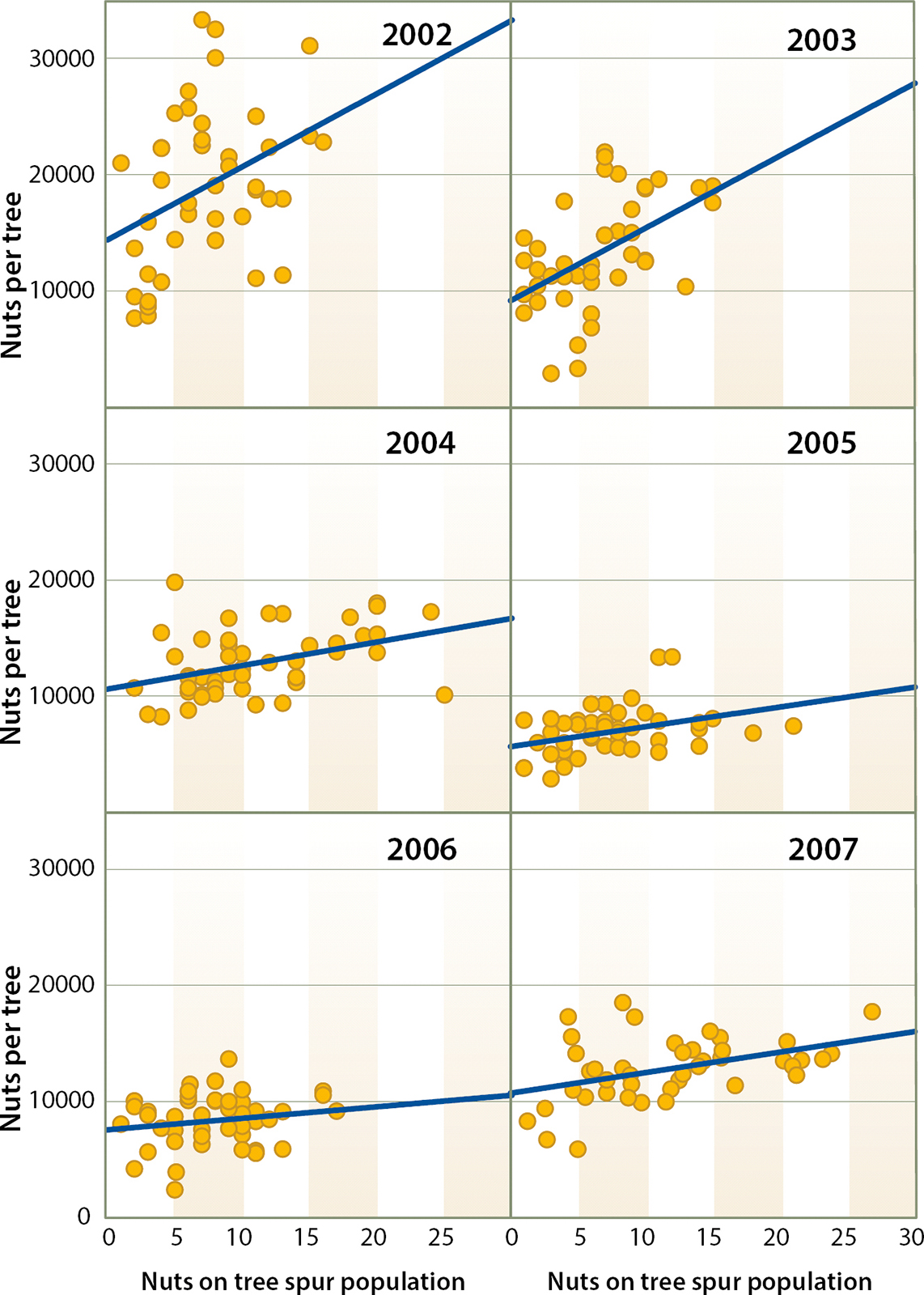
Fig. 5. Relationship between number of fruits per tree and number of nuts on a tree's tagged spur population in 2002 (R2 = 0.14, P < 0.05), 2003 (R2 = 0.25 P < 0.01), 2004 (R2 = 0.16, P < 0.01), 2005 (R2 = 0.13, P < 0.05), 2006 (R2 = 0.028, P = 0.26) and 2007 (R2 = 0.18, P < 0.01). Tree spur populations were composed of 50 spurs per tree.
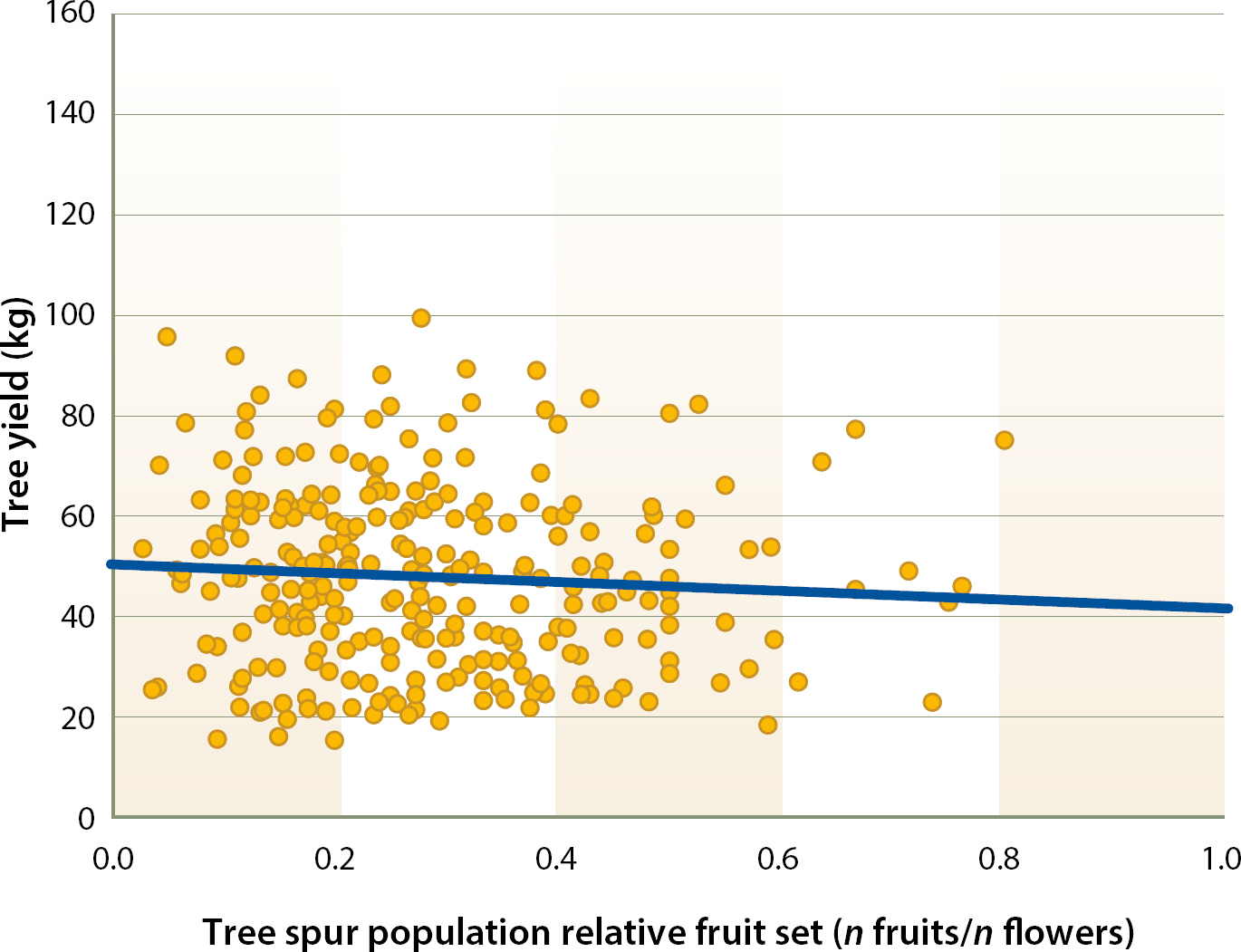
Fig. 6. Relationship between tree kernel yield and tree relative fruit set of the tagged spur population from 2002 to 2007 (R2 = 0.005 P = 0.23). 1 kg = 2.2 lb.
There were significant correlations between spur flower density and tree yield over years (fig. 7); for individual years, the relationship was significant in 4 of the 6 years of our experiment (table 2). On the other hand, the relationship between tree relative fruit set and tree yield was not significant in any of the 6 years of the experiment. However, it should be noted that the coefficients of determination (R2) were low due to the large number of points and the limited size of the spur sample compared with the total number of spurs borne by each tree; only 5.3% of the variability in tree yield can be explained by spur flower density.
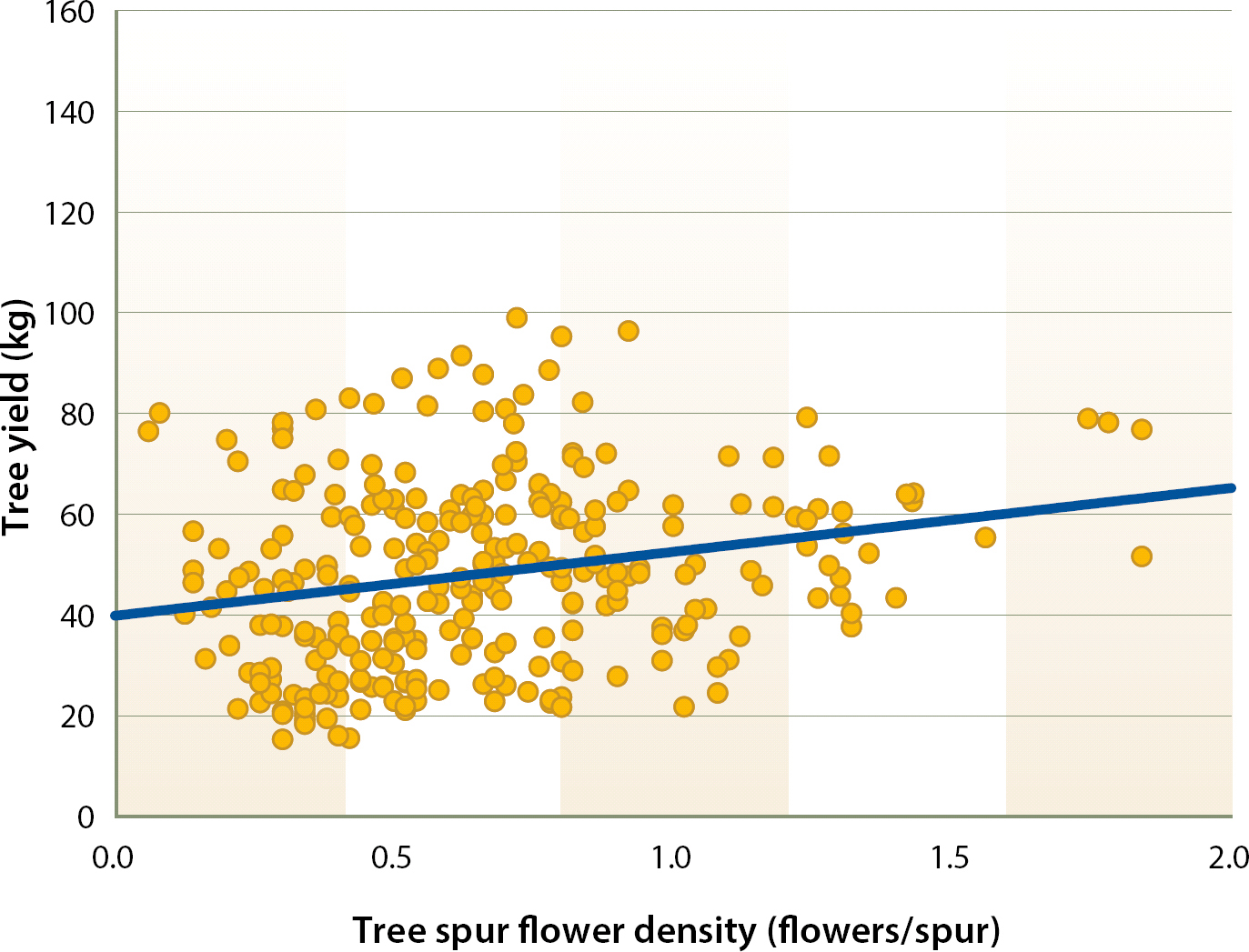
Fig. 7. Relationship between tree kernel yield and tree tagged spur flower density from 2002 to 2007. (R2 = 0.053 P < 0.0001). 1 kg = 2.2 lb.
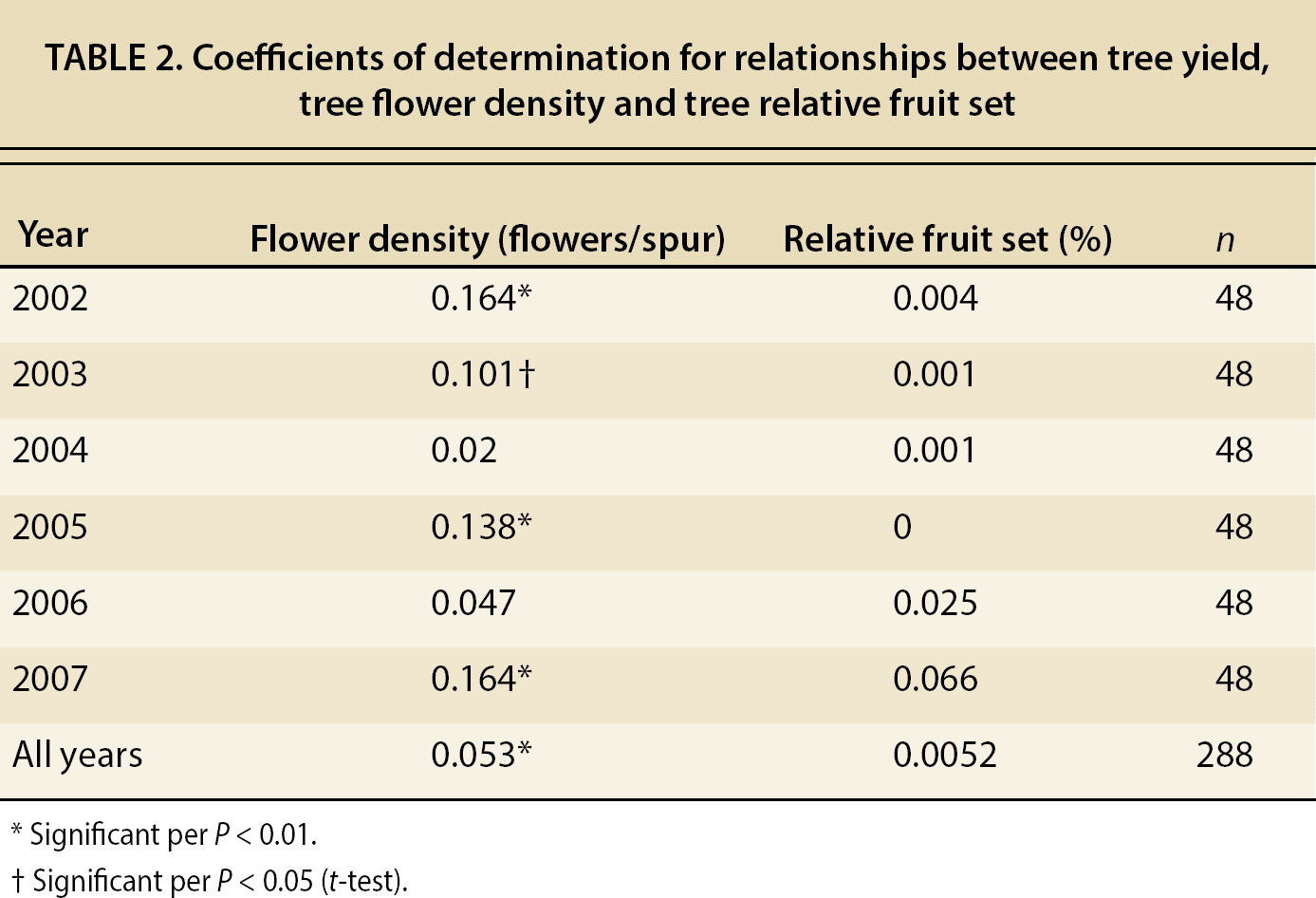
TABLE 2. Coefficients of determination for relationships between tree yield, tree flower density and tree relative fruit set
These results support the validity of flower density as an important parameter in the evaluation of almond cultivars (Kodad and Socias i Company 2008; Socias i Company et al. 1998). These data support the importance of total flower production for obtaining large crops.
Maximizing productive spurs
As a result of these spur dynamics studies, it is clear that the key to optimizing yields in commercial almond orchards is to focus on maximizing healthy populations of productive spurs.
Some spur mortality is unavoidable and linked to insufficient spur leaf area associated with spur bearing and spur shading (Lampinen et al. 2011; Tombesi et al 2011; Tombesi et al. 2015). Thus, continued productivity is dependent on spur renewal that is achieved by ensuring that there is annual growth of as many existing spurs as possible and new shoots that provide sites for new spurs (Esparza et al. 2001). Health of spurs is also a function of total canopy light interception and good light distribution with the tree canopy (Lampinen et al. 2011). It is clearly important to select cultivars with the ability to produce large numbers of flowers (Kodad and Socias i Company 2008) and have crop management practices (especially proper irrigation and fertilization) aimed at limiting abiotic stresses during the vegetative season (Esparza et al. 2001; Goldhamer and Smith 1995; Goldhamer and Viveros 2000).
In an experiment not potentially biased by experimental manipulation (i.e., deblossoming and hand pollination), these results support the assertion of Kester and Griggs (1959) that reductions in total number of flowers due to adverse orchard conditions are not likely to be compensated for by increased relative fruit set when adequate pollinizers and pollinators are present and can result in some measure of crop reduction. Such was the case in this study since it was conducted in an orchard in which the ‘Nonpareil’ trees were flanked by two pollinizer cultivars selected for bloom overlap with ‘Nonpareil’ and relatively high populations of bee pollinators were placed in the orchard each year to facilitate pollination. Had such factors not been present in the orchard during bloom, it is likely that relative fruit set would have varied even more among years and measured tree yields would have been more dependent on variations in relative fruit set.

Almond nuts and hulls at harvest time.
Yield in almond is related more to the abundance of flowers than the relative number of flowers that set fruit

