


All Issues
Hedgerows enhance beneficial insects on farms in California's Central Valley
Publication Information
California Agriculture 65(4):197-201. https://doi.org/10.3733/ca.v065n04p197
Published October 01, 2011
Abstract
Hedgerows of native California shrubs and perennial grasses bordering field crops were examined for the abundance of beneficial and pest insects compared with adjacent weedy areas. During 2 years of sampling in the Sacramento Valley, hedgerows attracted more beneficial than pest insects, while weedy areas showed the opposite trend, attracting significantly more pest than beneficial insects. We conclude that replacing weedy areas at field crop edges with managed hedgerow plantings will sustain or increase beneficial rather than pest insects on farms.
Full text
Hedgerows are rows of trees, shrubs, forbs and grasses that surround farm fields. They may be remnants of existing vegetation from cleared lands, a result of natural plant dispersal, or established via direct plantings (CAFF 2004;Long and Anderson 2010). Their many benefits include enhanced weed control, soil fauna, erosion control, sediment retention, game hunting, biodiversity, and air- and water-quality protection (Hannon and Sisk 2009; Kleijn et al. 2006; Kort et al. 1998; Smith et al. 2008). There also is evidence that hedgerows may increase the abundance of beneficial insects such as pollinators and natural enemies, possibly improving crop pollination and biological pest control in adjacent crops (Griffiths et al. 2007; Hopwood 2008; Thomas and Marshall 1999).
The enhanced biodiversity and potential ecosystem service benefits of hedgerows have prompted the USDA Natural Resource Conservation Service (NRCS) and Resource Conservation Districts to support growers in planting native shrubs and perennial grasses on their farms. Thirteen miles of hedgerows were established on California farms in 2009, compared to 3 miles in 2005. However, the adoption of hedgerows on farms is constrained by a lack of information about how they will alter pest and natural enemy communities in field edges as well as the benefits they may provide, including biocontrol of pests in adjacent crops (Brodt et al. 2009).

Hedgerows of California native shrubs and perennial grasses, including at Fong Farms in Yolo County, were compared to weedy field margins for the abundance of beneficial and pest insects.
The type of field edge habitat around farmlands influences the abundance and diversity of insects they attract, including pests that may be of concern to growers in adjacent crops (Pease and Zalom 2010). Our study evaluated how hedgerows of California native shrubs and perennial grasses affect beneficial and pest insect abundance in comparison to weedy field edges.
Hedgerows of shrubs and grasses
Beneficial and pest insects were examined in four hedgerows in Yolo County for 2 years.
Species planted.
Hedgerows at each site consisted of a row of perennial shrubs bordered by native perennial grasses. They ranged from 1,000 to 1,800 feet (305 to 550 meters) long and were established in 1996. Plant species composition for each site varied slightly, but all contained California lilac (Ceanothus griseus), coffeeberry (Rhamnus californica), California buckwheat (Eriogonum fasciculatum), toyon (Heteromeles arbutifolia), elderberry (Sambucus mexicana) and coyote brush (Baccharis pilularis). These are drought-tolerant native California shrubs that provide pollen and nectar for beneficial insects (Bugg et al. 1998; Long et al. 1998) and have successive and overlapping bloom periods (table 1).
The perennial grass stands were planted 10 feet (3 meters) wide along one or both sides of the shrubs to help suppress weeds and create overwintering habitat for natural enemies. The grasses included purple needlegrass (Nassella pulchra), nodding needlegrass (N. cernua), California melic (Melica californica), onesided bluegrass (Poa secunda), blue wildrye (Elymus glaucus) and creeping wildrye (Leymus triticoides).
All four hedgerows were adjacent to approximately 80 acres (32 hectares) of rotational field crops typical of crop production in this region, including wheat, processing tomatoes and alfalfa. At each hedgerow site, insect populations were monitored in an adjacent weedy, relatively unmanaged area (mowed or sprayed once or twice a year) of about 1,000 square feet. The primary herbaceous weeds in these adjoining weedy areas were wild mustard (Sinapis arvensis), black mustard (Brassica nigra), wild radish (Raphanus raphanistrum) and knotweed (Polygonum spp.).
Monitoring and identification.
Insects on hedgerow shrubs were monitored every 2 weeks from April to November 1999 and March to November 2000. At each sampling, two plants from each of the shrub species were randomly chosen within each hedgerow and visually inspected for insects. To assess the more mobile insect groups, such as syrphids, tachinids, lacewings and wasps, the number of visitors to each plant was observed and recorded for 2 minutes. Small insects that were not readily visible inside the flower heads were sampled by shaking all the flower heads on each shrub over a white sheet of paper and counting the number of insects dislodged. Weather conditions were monitored, and insects were sampled when temperatures were generally between 75°F to 85°F (25°C to 30°C) with sunny or bright overcast skies, and the fields were dry. In the early spring and fall samplings, temperatures were cooler and samples were taken as long as the temperature did not fall below 60°F (16°C).
Insects were identified to the taxonomic levels feasible from visual observation, by experienced observers who had carried out preliminary sampling in the hedgerows during 1997 and 1998. The UC Davis Bohart Entomology Museum also helped with species identification. The types and numbers of insects observed were recorded (table 2; fig. 1). The pest insects sampled were those of concern in adjacent field crops; the beneficial insects sampled were those that feed on major field crop pests. Few caterpillars (Lepidoptera), aphids, spider mites or leafminers were found in the hedgerows, so they were not included in our insect counts or data analyses. Thrips were not included because at the time of this study they were not considered a major field crop pest in this region. However, due to the introduction of new thrips-transmitted viruses since this study, our current research is focusing on monitoring thrips in hedgerows.


TABLE 2. Beneficial and pest insects sampled in hedgerows of native shrubs and perennial grasses and in weedy areas, Yolo County
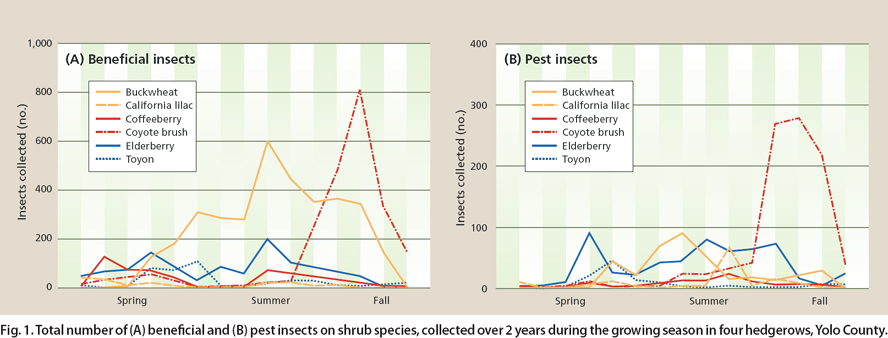
Fig. 1. Total number of (A) beneficial and (B) pest insects on shrub species, collected over 2 years during the growing season in four hedgerows, Yolo County.
Plant size and sampling frequency.
To standardize the counts from visual observations and flower shake samples among plants, the size of each shrub sampled was estimated by measuring the average length and width of each plant (most plants were relatively circular) multiplied by height, giving an approximate square area in meters. Insect numbers were divided by plant size (which varied considerably, particularly in height), providing a measurement of insect abundance per square meter.
The perennial grasses in the hedgerows and adjacent weedy areas were also sampled every 2 weeks from April to November 1999 and March to November 2000 using a standard sweep net (UC IPM 2006). At each site, 10 sweeps were taken in each of four different areas of both the hedgerow grasses and weedy areas.
The abundance of beneficial insects was consistently greater than pests in the hedgerow shrubs compared with weedy areas during each season.
Statistical analysis. Beneficial and pest insect abundances within each vegetation type (shrub, grass and weed) were compared from spring to fall using a full factorial mixed model ANOVA (SAS 1999). Sample period was a repeated factor; insect type (pest or beneficial), year (1 and 2) and plant species (1 to 6) were fixed effects; site (1 to 4) was a random factor; and abundance of insects was the response variable.
For post hoc comparisons, we coded sample period by season (spring, summer and fall) and included season and its interactions as fixed effects. Abundance data were Poisson-distributed and square-root (plus constant of one) transformed before analyses. To compare shrub data (collected by surveys of plants) to grass and weed data (collected by standardized sweep samples), we compared the proportion of beneficial insects among vegetation types using a general linear model with a binary distribution and a logit link function (SAS 1999).
Insect population counts
Beneficial insects.
Of 8,045 beneficial insects collected in the four hedgerows over 2 years, 31% were minute pirate bugs, 17% syrphid flies, 13% assassin bugs, 13% tachinid flies, 10% big-eyed bugs, 6% lacewings, 6% wasps, 3% lady beetles and 0.4% damsel bugs. The greatest abundance of beneficial insects was collected on California buckwheat, followed, in decreasing amounts, by coyote brush, elderberry, coffeeberry, toyon and California lilac. The greatest beneficial insect abundance on each shrub species coincided with the bloom period of that species, when nectar and/or pollen were available (fig. 1A).
Pest species.
Of 2,278 pests collected in the four hedgerows over 2 years, 42% were spotted cucumber beetles, 25% Lygus bugs, 18% flea beetles and 14% stink bugs, with the greatest abundance also occurring during plant bloom (fig. 1B). The increased number of pests on California buckwheat during summer was caused primarily by Lygus bugs. Similarly, spotted cucumber beetles were the primary cause of the pest population increase in coyote brush during the fall bloom. Flea beetles were most numerous on the hedgerow plants during summer, and stink bugs were most abundant during summer and fall, when the shrub berries were ripening.
Insect abundance.
Of 10,323 total insects collected in the hedgerows during the growing seasons over 2 years, 78% were beneficial insects and 22% were pests. The abundance of beneficial insects was consistently greater than pests in the hedgerow shrubs compared to weedy areas during each season (P < 0.0001 Bonferroni; fig. 2). Overall, a greater abundance of insects was collected in year one than year two (F1,11.3 = 7.92, P = 0.0164). But there was no difference in relative abundances of pest and beneficial insects between the two years (year by insect type interaction; F2,1252 = 0.01, P = 0.940) or interaction among year, insect type and season (F2,1252 = 2.18, P = 0.114).
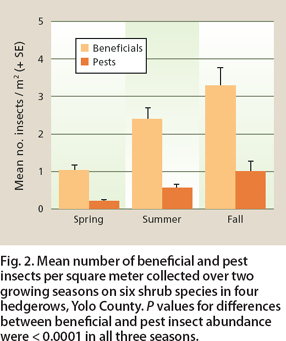
Fig. 2. Mean number of beneficial and pest insects per square meter collected over two growing seasons on six shrub species in four hedgerows, Yolo County. P values for differences between beneficial and pest insect abundance were < 0.0001 in all three seasons.
Examination of sweep sample collections showed that pests were more abundant in the weeds than in the native perennial grass stands in spring (t317 = −6.17, P < 0.0001 Bonferroni), summer (t317 = −13.20, P < 0.0001 Bonferroni) and fall (t317 = −5.32, P < 0.0001 Bonferroni; fig. 3). Beneficial insect abundance increased on weeds during summer but not to the same extent as the pest insects. In summer, the grasses dried and few insects were found.
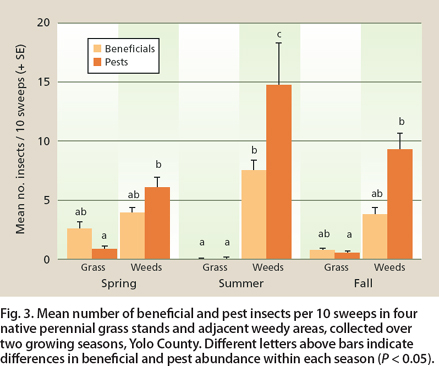
Fig. 3. Mean number of beneficial and pest insects per 10 sweeps in four native perennial grass stands and adjacent weedy areas, collected over two growing seasons, Yolo County. Different letters above bars indicate differences in beneficial and pest abundance within each season (P < 0.05).

In this study, hedgerows enhanced the ratio of beneficial to pest insects compared with weedy areas. Plantings at Sierra Orchards in Solano County include deer grass, California lilac and elderberry. Inset left to right, the beneficial insects identified included lady beetles, syrphid flies and their larvae (feeding on aphids).
Across seasons (χ21 = 384.11, P < 0.0001) and within each season, there was a greater proportion of beneficial to total (beneficial plus pest) insects in shrubs than in weeds, with grasses having a proportion of beneficial to pest insects intermediate to shrubs and weeds (fig. 4).
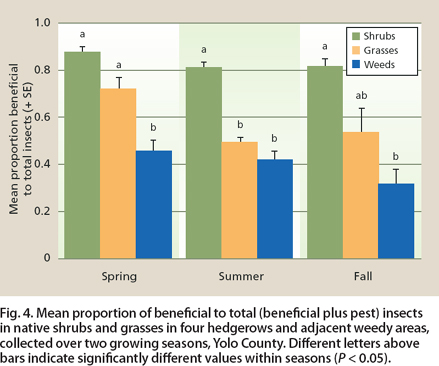
Fig. 4. Mean proportion of beneficial to total (beneficial plus pest) insects in native shrubs and grasses in four hedgerows and adjacent weedy areas, collected over two growing seasons, Yolo County. Different letters above bars indicate significantly different values within seasons (P < 0.05).
Growing interest in hedgerows
Our results show that field edge plantings of native California shrubs and perennial grasses can enhance beneficial insect abundance. The enhancement of beneficial insects may occur in several ways. First, most beneficial insects require or benefit from nectar or pollen sources from flowering plants that hedgerows provide, helping them survive and reproduce, especially during times of prey scarcity (Bugg et al. 1998). This was apparent in our study; beneficial insect abundance was greatest on shrubs during bloom, suggesting that insects were using floral resources. Second, hedgerows provide some beneficial insects with alternative prey or hosts, which may also be most important during wintertime (Corbett and Rosenheim 1996). Third, hedgerows provide beneficial insects with overwintering habitat, which is important when neighboring fields are cultivated and fallow for the winter, and there are few other refuges (Dennis et al. 1994).
Our study provides evidence that hedgerow plantings can enhance ratios of beneficial to pest insects compared with weedy areas, where pests were found in significantly greater abundance than beneficial insects. The extent to which this enhanced abundance of beneficial insects in hedgerows will improve biological pest control in adjacent crops is largely unknown. Previous research showed that beneficial insects used floral resources provided by hedgerows and moved into adjacent crops (Long et al. 1998). In a review of natural pest control, 74% of cases studied showed that landscapes with high proportions of noncrop habitat had enhanced natural enemy populations in crop fields (Bianchi et al. 2006). Further, eliminating edge weeds (by mowing or spraying) or replacing them with managed vegetation such as native perennial grasses has led to reduced pest pressure in adjacent crops (Ehler 2000; Mueller et al. 2005; Pease and Zalom 2010).

Compared with weedy areas, hedgerows did not increase populations of insects such as redshouldered, consperse and southern green stink bugs, pests of tomatoes and other crops.

Current studies are examining insect populations in crops adjacent to hedgerows and weedy field edges, such as, above, black mustard near an agricultural field.
For improved biocontrol through hedgerow plantings on farms, it is important that plants enhance beneficial insects without increasing pest populations (Fiedler and Landis 2007). In our study, the native shrubs and perennial grasses, though used by pests, were not as preferred as the weeds were, as noted by the significantly greater proportion of beneficial insects compared with pests in the hedgerow plantings. Although California buckwheat attracted Lygus bugs during summer and coyote brush attracted spotted cucumber beetles during fall, beneficial insect abundance was far greater than pests on those plants.
As noted earlier, one of the impediments to growers adopting hedgerows is the concern that they will harbor and enhance pest insect populations in adjacent crops. Our data show that hedgerow plantings can sustain or enhance beneficial insects and serve as replacement vegetation for weedy field edges, which harbor pests.
Recently, more hedgerows have been adopted in the Sacramento Valley. In our current studies, we are standardizing the crop adjacent to hedgerows and examining insect populations and pest control in the crop. These studies will address the question of whether hedgerows are concentrating existing populations of beneficial insects or whether they are increasing beneficial populations for enhanced pest control in adjacent crops. The study reported here and our current evaluations of the economic benefits of hedgerows on pest control may lead to the wider adoption of hedgerow plantings on farms, helping to enhance the many ecosystem service benefits they provide in agricultural landscapes.
Citations
Hedgerows enhance beneficial insects on farms in California's Central Valley

