Calag Archive
Calag Archive
Management strategies outlined: Research reveals pattern of cucurbit virus spread
Publication Information
California Agriculture 46(2):35-40.
Published March 01, 1992
PDF | Citation | Permissions
Abstract
Diseases caused by aphid-vectored viruses result in severe economic loss to Southern California melon growers. Information gathered over the past several years has given researchers new directions for managing this production problem.
Full text
![Green peach aphid (Myzus persicae [Sulzer]), a common species in the Imperial Valley which vectors both WMV2 and ZYMV.](http://ucanr.edu/sites/calagjournal/archive/?image=img4602p35.jpg)
Green peach aphid (Myzus persicae [Sulzer]), a common species in the Imperial Valley which vectors both WMV2 and ZYMV.
Spring cucurbits in the southern desert valleys of California and Arizona have been plagued by diseases caused by aphid-vectored viruses for many years. These diseases, which are characterized by foliar mosaic symptoms, were studied initially in the mid-1940s and have been identified as cucumber mosaic virus (CMV), papaya ringspot virus-type W (PRSV), watermelon mosaic virus 2 (WMV2), and zucchini yellow mosaic virus (ZYMV). Our extensive surveys in spring-grown melons throughout the Imperial Valley over the past 5 years have identified WMV2 and ZYMV as the most common viruses that lead to the majority of crop loss.
We have attempted several virus disease control measures in melons in the Imperial Valley. Insecticide and oil applications have not proved effective for reducing disease incidence. We suspect this failure is due to the large number of winged aphids that land on the plants, the inability to maintain complete coverage between applications and the rapidity with which virus transmission occurs. Row covers and/or reflective plastic mulches reduce the rate of virus spread by preventing aphid access (covers) or repelling winged aphids from landing on the plants (mulches), but presently these techniques are prohibitively expensive. Therefore, we have devised management strategies of diseases in the Imperial Valley caused by WMV2 and ZYMV based on (1) the transmission characteristics of each virus and their relationships to aphid vectors, (2) the response of cucurbit plants to infection and (3) the host ranges of the two viruses.
Transmission characteristics
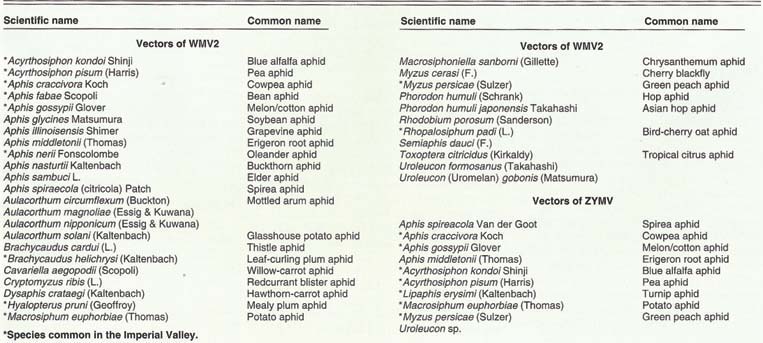
Both viruses are members of the poty-virus group and are transmitted from plant to plant by aphid vectors. Although it is possible to transmit these viruses mechanically by rubbing the leaves of a healthy plant with sap from an infected plant, the majority of transmission in the field is by aphid vectors. Our research, coupled with other studies, has shown that at least 35 aphid species transmit WMV2 and at least 9 species transmit ZYMV (table 1).
Both viruses are transmitted in a non-persistent fashion, which has the following implications. First, only a short feeding time is necessary for an aphid to acquire virus from an infected plant. (We have conducted transmissions in the field with acquisition times of less than 15 seconds.) Second, there is no time period required between acquisition and transmission; an aphid can transmit virus immediately to a healthy plant. Third, the aphid can infect a healthy plant with a short inoculation probe, once again in as little as 15 seconds. These brief acquisition and inoculation times limit the usefulness of aphicides to reduce the spread of WMV2 and ZYMV, because it generally requires longer than 15 seconds for aphids to obtain a lethal dose of the aphicide. Finally, aphids retain these viruses for a short time; we have determined that aphids which have acquired ZYMV lose the ability to transmit in less than 4 hours at 70°F, although longer retention times have been determined at cooler temperatures.
Plant response
Both viruses cause mosaic symptoms on the cantaloupe foliage ( see photograph, p. 40 ) and generally result in more yield loss when plants are inoculated early in the growth Cycle, Qur Studies indicate that if a plant is infected with ZYMV before setting fruit, the plant will not produce any fruit. If infection occurs during early fruit set, the plant may abort the existing fruit or the fruit may become deformed and unmarketable ( see photograph, p. 40 ). WMV2, on the other hand, does not cause this severe fruit reaction. Plants infected with WMV2 show mosaic on the foliage, but will continue to produce fruit unless infection occurs in the seedling stage. Plants infected after fruit are set will continue to fill fruit and produce normal melons.
Host ranges
Another substantial difference between ZYMV and WMV2 is in the host ranges of the viruses (table 2). From these lists, it is apparent that WMV2 has the potential to be in numerous wild and cultivated plants through the winter months in the Imperial Valley, whereas the sources for ZYMV are limited. In the Imperial Valley, the only common plants known to harbor ZYMV are in the Cucurbitaceae. The other known ZYMV hosts, which consist of henbit, fenugreek, crowfoot and the ornamental, bluewings, are uncommon, if present at all, in the valley. Since cucurbits are susceptible to frost, we suspect that the abundance of potential ZYMV host is nominal after a cold winter in which the valley experiences freezes.
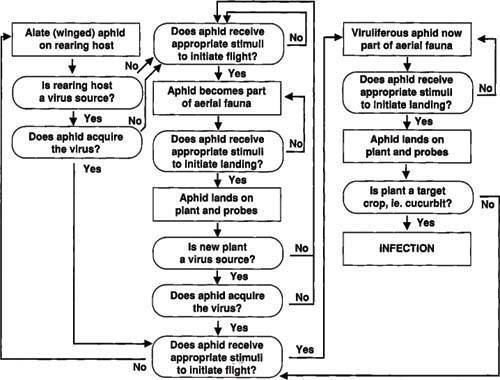
Fig. 1. Model of steps involved for aphid acquisition of a non-persistently transmitted virus and transmission to healthy plants.
Development of epidemics
For any plant to become infected, a defined pattern of events must take place sequentially (fig. 1). First, an aphid of a species that is capable of transmitting virus must be present; not all aphid species will vector all viruses. A winged form must receive stimuli causing it to leave the plant on which is was born, which may or may not be the plant from which it acquires virus. Examples of stimuli that might cause aphid migration include declining host conditions (caused by drought, freezing or natural plant senescence), crowding or escape from predators.
If the aphid is not viruliferous (carrying the virus), it must fly to and land on an infected plant. The aphid must probe the virus source plant, and if its stylets contact a cell with transmittable virus, it will become viruliferous. At this time, another set of stimuli is required to cause the aphid to leave the virus source plant. If the aphid colonizes the plant, the epidemiological cycle will be interrupted until the aphid, or its offspring, receives stimuli to leave the source plant.
We suspect that most frequently the virus source plant is not an acceptable colonizing host for the aphid; therefore the aphid spends only enough time on the source plant to sample the plant, during which it acquires virus. After leaving the source plant, the aphid must find a healthy cucurbit plant, alight on this plant and probe. As noted, an aphid which has acquired virus is able to infect a healthy plant for only a short time (a period of hours) under normal cucurbit growing temperatures.
Knowing this series of defined events and knowing that each has to happen in the proper order, one would think that the probability of a plant becoming infected is low. However, estimated melon yield loss to ZYMV and WMV2 in the 1990 Imperial Valley spring crop was in excess of $22 million. We believe that epidemics of this proportion occur only when the number of available virus source plants or the density of winged aphids is high. With this as our working model, we began research in 1985 to identify weak points in the epidemiological cycle that could be exploited in an integrated virus management program.
Research
In the spring of 1985, we observed that mosaic diseases caused widespread yield loss. Many fields were disced before harvest, which suggested that ZYMV was common in the Imperial Valley. Records from the winter of 1984–85 showed that there were only 2 days in which temperatures were below 30°F; cucurbit hosts of ZYMV were probably abundant in the valley when melon plants were emerging in commercial fields.
![Plants known to serve as hosts for WMV2 and ZYMV. (Plants found in the Imperial Valley are designated by an asterisk [∗].)](http://ucanr.edu/sites/calagjournal/archive/?image=tab4602p37.jpg)
TABLE 2. Plants known to serve as hosts for WMV2 and ZYMV. (Plants found in the Imperial Valley are designated by an asterisk [∗].)

Fig. 2. Fields surveyed in the Imperial Valley in 1989 which showed early infection with ZYMV2 and ZYMV.
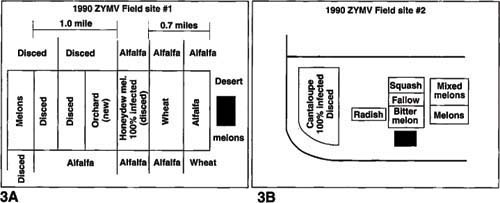
Fig. 3. Fields surveyed in the Imperial Valley in 1990 which showed early infection with ZYMV. Locations of ZYMV sources are indicated by the dark areas
In 1986, disease severity varied throughout the valley so we obtained field locations (identified by irrigation canal and gate) from the Imperial County Agricultural Commissioner's Office and visited each melon field. Conducting a single survey of the 273 melon fields near harvest, we visually rated each as having high, moderate or low disease incidence. This survey showed that most of the severely infected fields were in the southern half of the valley, an indication that virus may have come from plants growing in the Mexicali Valley of Mexico.
Cooperating with the Department of Geography at UCR, we digitized the irrigation system of the Imperial Valley (nearly 5,500 individual field locations) into a computer data file, so that melon field lists from the agricultural commissioner could be mapped readily. The Geographic Information System (GIS) mapping software package, “ARC-INFO,” allowed us not only to accurately map all fields in the valley, but to spatially analyze information about each field that was gathered on surveys. In this way we described the spatial and temporal spread of disease throughout the 350,000-acre agricultural area in 1988 and 1989.
At each field we randomly selected 100 plants each from the northwestern and southeastern corners, visually assessing the percentage of plants infected with mosaic-causing viruses. In each field, we collected samples from plants with mosaic symptoms to analyze for ZYMV, WMV2, CMV and PRSV, using an enzyme-linked immunosorbent assay (ELBA). Two surveys were conducted each year, one when fields were near the perfect flower growth stage and one just before harvest. After surveying 227 fields in 1988 and 215 fields in 1989, we found that 87% and 75% of the fields, respectively, had WMV2, ZYMV or a combination of these two viruses. CMV and PRSV rarely were found.
During our early survey in 1989, we found four fields that had disease gradients from one side of the field to the other (fig. 2). Based on samples analyzed by ELISA, we determined that two of the fields were infected predominantly with WMV2 (fig. 2 a, b) and two were infected primarily with ZYMV (fig. 2 c, d). We conducted more extensive sampling in these fields to resolve the spatial pattern of virus spread.
In the first WMV2 field (fig. 2a) we determined that the gradient of virus was from the southeastern corner to the northwestern corner. To the southeast of the melon field, large numbers of green peach aphids were infesting a field of carrots. The disease pattern suggested that (1) WMV2 was introduced into the field from one of the many WMV2 host sources and (2) the high density of aphids from the carrots moving across the melon field resulted in the WMV2 gradient that we observed.
In the second WMV2 field (fig. 2b), incidence was heaviest on the western side of the field, while the eastern side was less infected. Again we surveyed the surrounding vegetation and found that tomatoes and sudangrass, both potential aphid sources, were to the west of the field. The tomato field had an infestation of potato aphids, another known vector of WMV2. In both WMV2 situations, we felt that it might have been possible to reduce the rate of virus spread had the melon producer been able to reduce aphid densities in the surrounding fields.
The two fields with ZYMV showed a different pattern of disease incidence, which we determined was consistent with spread from a local point source. In the first ZYMV field (fig. 2c), there was a gradient of virus from south to north. Further searches revealed a garden planting of squash that was infected with ZYMV (the squash was located near the dark area in fig. 2c). The second ZYMV field (fig. 2d) had a disease gradient from north to south, and once again the dark region represented a residential area. Although we were prevented from surveying the residential area, we suspect that protected cucurbit plants were on the premises.
The information gathered from these four fields, coupled with our observations from 1986-1989, allowed us to formulate the following hypothesis concerning spring virus epidemics in the Imperial Valley. First, we think that most epidemics originate locally; long-range immigration of viruliferous aphids is not frequent. Second, after cold winters the only surviving source plants for ZYMV are in areas that protect them from cold weather. These are in home garden plantings of squash or sponge gourd or commercial plantings of melons or squash grown under plastic or in greenhouses. The number of surviving WMV2 hosts, on the other hand, is not as likely to be affected by low temperatures. Thus, for the less pathogenic WMV2, severe epidemics will be caused by the presence of nearby aphid source plants very early in the melon-growing season.
Testing the hypothesis
The spring 1990 season provided us an opportunity to examine our hypothesis. The winter conditions were the coldest in recent history (9 days below 30°F), and we predicted a light ZYMV year. We suspected that the valley would be devoid of all ZYMV source plants, and the overwintering survival of ZYMV would be negligible. However, in early April, we observed two fields that were devastated by virus disease; both fields were disced without harvesting a single fruit. From one of the fields (fig. 3a), we collected 100 plant samples and determined, by ELISA, that all but 1 of the samples had ZYMV. Surveying the surrounding area, we found a planting of an unknown squash variety that had been planted in the fall of 1989 and grown in a house covered with plastic, preventing the plants from freezing (dark area in fig. 3a).
At the time of our visit to the plastic house, the plants were nearly dead, yet leaf samples were positive for ZYMV. There was also evidence (curled leaves, residues of honeydew and sootymold, and a remnant aphid population) of high melon aphid pressure on the squash. We were informed that as conditions in the house became warm, the grower removed roof sections of plastic to provide ventilation. This could have served as an escape for aphids leaving the squash plants. The rapidity with which the nearby melon field became infected (0 to 100% in 10 days) suggested that a heavy flight of viru-liferous aphids landed in the field over a short time period.

ZYMV-infected yellow squash
From the other field (fig. 3b), we collected 15 plant samples, finding 11 with ZYMV. Surveying this area, we found a volunteer planting of sponge gourd, a known ZYMV host (table 2), growing in an area protected from frost at a nearby residence. Once again at the time of the survey, we collected leaves from the sponge gourd, which tested positive for ZYMV. A compounding factor in this area was the presence of a radish field that was replete with green peach aphids. Further research indicated that virus from the ZYMV infections in these two fields spread to neighboring fields resulting in the low average yields of 1990 (estimated at 200 cartons per acre compared to normal production of 500 to 550 cartons per acre).
Management strategies
The primary focus of our proposed integrated virus management program is to limit ZYMV source plants by conducting a thorough survey of areas surrounding melon fields before melon emergence. First, particular attention should be paid to residential areas that may have gardens protected from the cold or commercial cucurbit plantings grown under plastic. Once the locations of these plants are identified, ZYMV infection needs to be determined. Once the plants are infected with ZYMV, they should be removed because there is little chance they will produce a substantial yield.
Second, growers should be keenly aware of surrounding crops and weeds that may serve as aphid sources. Nearby weeds should be removed, and neighboring crops should be observed for aphid densities. Although many winter-spring crops harbor vector aphid species, we found particularly severe infestations in weedy cole and lettuce crops abandoned after harvest in the early spring. Cucurbit fields near these abandoned fields probably will have severe ZYMV and/or WMV2 disease problems.
Producers commonly inquire about the use of aphicides to prevent or slow the spread of ZYMV and WMV2 in their fields. This tactic has not been shown to produce a profitable return because of the rapidity with which transmission occurs. Even if a lethal dose of pesticide were acquired by an aphid during feeding, the virus already would be in the plant because it can be deposited in as little as 15 seconds. However, aphicide applications to surrounding crops that are serving as aphid sources may be helpful.
We suggest that the grower develop an awareness of the sources of virus inoculum around his field and the vector pressure that is responsible for carrying virus into his field. Once these are determined, the producer then can concentrate on reducing both parameters so that the probability for growing healthy cucurbits is increased.

At left and above, normal cantaloupe and watermelon are compared with diseased fruit. Small and deformed fruit are ZYMV-infected. Below left, healthy cantaloupe; at right, mosaic symptom on cantaloupe foliage characteristic of infection with ZYMV or WMV2.
Management strategies outlined: Research reveals pattern of cucurbit virus spread

