Calag Archive
Calag Archive
Livina mulches suppress aphids in broccoli
Publication Information
California Agriculture 48(4):24-28.
Published July 01, 1994
PDF | Citation | Permissions
Abstract
A living mulch combines an annual crop with a cover crop to provide pest control and soil protection. Results of a study in the Salinas Valley show cabbage aphid infestation can be lowered on broccoli grown in living mulches compared to clean cultivation, possibly because light reflectance patterns are less attractive to incoming aphids. Early and intense mowing of the cover crop and adequate irrigation maintained broccoli yield in living mulches relative to clean cultivation. Lack of specialized equipment is the main limitation to the commercial application of living mulches at this time.
Full text

At left, transplanting broccoli into cover crop.

Below, broccoli transplants are placed into a 4-inch-wide cultivated band in the cover crop.
A living mulch — an interplanting system in which a low-growing grass or legume is grown with an annual crop — can be an effective alternative to pesticides in broccoli fields and, if properly maintained, does not reduce crop yields. As increasing restrictions are placed on the sale and use of pesticides, growers are seeking alternatives, especially in pest-sensitive crops such as broccoli. In addition to replacing pesticides, living mulches smother weed growth, protect the soil from wind and water erosion, enhance soil fertility and structure, and reduce soil compaction from heavy equipment. They can play a role in conventional as well as organic agriculture.
In 1991, 88,000 acres of broccoli were under cultivation in California, mainly in the Salinas Valley, with a value of $215 million. On broccoli, the cabbage aphid, Brevicoryne brassicae, can reach economically damaging levels by moving into the developing flower buds and rendering the head unmarketable. Because broccoli heads have virtually no tolerance for this aphid, growers may spray their broccoli fields four to six times during periods of highest aphid activity.
To become well established and suppress weed growth, the cover crop that functions as the living mulch should have vigorous early growth, the ability to regrow effectively after mowing and a dense, mat-like growth habit. Unfortunately, these are the very characteristics that will result in competition with the main crop for resources such as light, water and nutrients, potentially causing unacceptable yield declines in the main crop. As a consequence, most living-mulch research has entailed suppressing the cover crop during the main crop's growing season by applying sublethal concentrations of herbicides.
Our studies were designed to determine whether living mulches could be used in place of insecticides to protect broccoli from cabbage aphid infestation. By measuring reflected light from the crop, nitrogen levels and aphid parasitism, we hoped to understand the mechanisms involved. We were also interested in determining whether yields could be maintained (relative to clean-cultivated broccoli) by substituting mechanical suppression of the cover crop for chemical suppression.

At right, broccoli/white clover living mulch, 28 days after planting, fall 1990. The clean cultivated control can be seen in the background.
Experimental design
We conducted experiments at the Rural Development Center in the Salinas Valley in fall 1990 and summer 1991. The soil at the site is a Hanford sandy clay loam. The experiment was a 4 by 2 factorial, in a randomized complete block, split plot design. Blocks consisted of four 33-by-33-ft (10-by-10-m) plots, which were replicated four times. Three cover crops and a clean-cultivated (no cover crop) control were the main plot variables and were randomly assigned to plots within blocks. The cover crops were perennial legumes (Rhizobium coated seed) seeded in September 1989 at the equivalent rate of 140 lb/ac (140 kg/ha), without fertilization. The legumes were white clover (Trifolium repens L.), strawberry clover (Trifolium fragiferum L. ‘O' Conners‘) and birdsfoot trefoil (Lotus corniculatus L. ‘Kalo’). The trefoil seed lot was infested with red clover (Trifolium praetense L.), a biennial that proved impossible to eliminate by mowing; together they formed a trefoil/red clover cover crop. We used different legumes to evaluate their competitiveness with the broccoli. The subplot variables consisted of either synthetic fertilizer or chicken manure and rice hull compost. Alleyways between plots were 13 ft (4 m) wide and were kept free of vegetation during the experiments with a hand-held rototiller.
Two weeks before each experiment, 4-inch (10-cm) rows were cultivated into each main plot on 24-inch (60-cm) centers. These spacings provided 83% soil cover in the living mulch plots. Five-week-old broccoli starts (‘Shogun’ in fall 1990 and ‘Arcadia’ in summer 1991) were placed 8.5 inches (22 cm) apart within the row, the equivalent of 30,000 plants per acre. Cover crops were mowed to prevent excessive shading of the young broccoli plants, 7 and 21 days after transplanting in fall 1990 and 7 and 14 days afterward in summer 1991. All plots were sprinkle-irrigated twice a week, at the rate of 1 acre-inch per irrigation.
Fertilizer was applied to the broccoli rows in each plot; the row middles were left unfertilized. The rates of compost used were those recommended by the manufacturer, and the rates of synthetic fertilizer were equivalent to those used by a local commercial broccoli producer. Compost (1.55–2.84–2.22 [N-P-K] at 20% moisture) was applied before planting at 32 cubic yd/ac (equivalent to 11 t/ac), representing an approximate application of 170 lb N/ac. Synthetic fertilizer (15-15-15 preplant and 20-0-0 postplant) was applied at rates of 200 lb N/ac for fall 1990 and 220 lb N/ac for summer 1991. Commercial broccoli growers in the Salinas Valley apply approximately 200 lb N/ac.
Fifty individual leaves and two broccoli plants were randomly selected from each subplot 35, 49 and 63 days after planting in fall 1990 and 22, 32, 42, 52 and 62 days after planting in summer 1991. The leaves were shaken to remove aphids, and parasitized aphids (mummies) were removed by hand. All leaves were removed and measured with a leaf area meter. In summer 1991, yellow aluminum pans filled with water were placed in each subplot to trap incoming aphids. Also, broccoli leaf samples were taken from the strawberry clover living mulch and from the clean-cultivated control 32, 42 and 52 days after planting, dried at 140°F (60°C) for 1 week, and then ground, screened and sent to the UC Division of Agriculture and Natural Resources analytical lab for nitrate-nitrogen analysis.
To measure light reflected from the broccoli plants, we converted a light meter to a spectral radiometer by covering the sensor with a series of colored glass filters. These filters split total reflected light into spectral wavelength bands (wavebands) of blue (400–455 nanometers [nm]), blue-green (455–515 nm), green (515–550 nm), yellow-orange (550–610 nm, fall 1990) or yellow (550–590 nm, summer 1991), and orange-red (610–700 nm, fall 1990) or orange (590–640 nm, summer 1991). The sensor was held 1 ft (30 cm) above three randomly selected broccoli plants in each subplot; measurements were taken 28, 42, 56 and 70 days after planting in fall 1990 and 22, 32 and 52 days after planting in summer 1991.
At harvest, 10 broccoli heads per subplot (20 in summer 1991) were cut 8 inches (20 cm) in length, and each head was carefully inspected for colonies of cabbage aphid. If at least one colony was found, the head was scored as infested and considered unmarketable. All treatment effects were determined by analysis of variance (ANOVA). Nonlinear regression analysis was used to determine the relationship between alate numbers and light reflectance intensity.

Fig. 1. Percent of harvested broccoli heads infested with cabbage aphid by cover crop and fertilizer type. ** = Mean of clean cultivation greater than living mulches in subplots given compost (P<0.01). †† = Mean of clean cultivation greater than living mulches.
Compost provides low nitrogen
Leaf nitrate levels of broccoli leaves from the 1991 season were significantly higher with synthetic fertilization than with compost on every sampling date (table 1), suggesting that nitrogen from the compost was not being made available to the broccoli at a fast enough rate. Nitrate levels were so disparate that the synthetic fertilizer treatment could be considered a high nitrogen application and the compost a low nitrogen one. Differences were 20-fold on 32 days, 9-fold on 42 days and 12-fold on 52 days after planting. No significant leaf nitrate differences were detected between the strawberry clover living mulch and the clean cultivation, and there was no interaction between cover crop and fertilizer factors.
![Mean whole-plant nitrate-nitrogen content of broccoli leaves from strawberry clover living mulch and clean cultivation by compost and synthetic fertilizer, summer 1991 (Parts per million [ppm] are rounded to the nearest 100)](http://ucanr.edu/sites/calagjournal/archive/?image=tab4804p26.jpg)
TABLE 1. Mean whole-plant nitrate-nitrogen content of broccoli leaves from strawberry clover living mulch and clean cultivation by compost and synthetic fertilizer, summer 1991 (Parts per million [ppm] are rounded to the nearest 100)
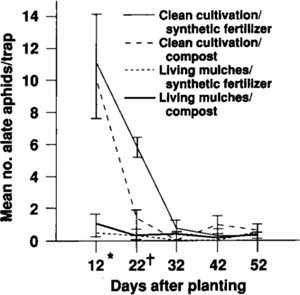
Fig. 2. Mean numbers of alate (winged) cabbage aphids captured in yellow water traps, summer 1991. * = Mean of clean cultivation higher than living mulches (P,0.001). † = Mean of clean cultivation/synthetic higher than living mulches/synthetic (P<0.01).
Differences were minor in all other measured parameters among the white clover, strawberry clover and trefoil/red clover levels of the cover crop factor; in the rest of this article we refer to these cover crops collectively as the “living mulches.”
Fewer early-season aphids
In each season, aphid infestation of harvested broccoli heads was affected by the interaction of cover crop and fertilizer factors. In fall 1990, infestation rates differed between living mulches and clean cultivation in compost subplots (7.5% in living mulch/compost compared with 55% in clean cultivation/compost); no significant difference in infestation was found between living mulches and clean cultivation in synthetic fertilizer subplots (the average was 8.8%). In summer 1991, infestation was higher in clean cultivation than in living mulch subplots at both fertilizer levels, but it was more severe in compost (40% in clean cultivation/compost and 10% in living mulch/compost) than in synthetic fertilizer subplots (15% in clean cultivation/synthetic fertilizer and 4% in living mulch/synthetic fertilizer). These findings are shown in figure 1.
In the 1990 experiment, early-season numbers of alate aphids (the winged, dispersing form of the aphid) per sample were many times lower on broccoli leaves from the living mulches (mean = 0.08 ± 0.05) than on leaves from clean cultivation (mean = 1.37 ± 0.27; F = 11.6, P = 0.002), but by the last sampling date no differences remained (living mulch mean = 0.20 ± 0.10; clean cultivated mean = 0.04 ± 0.04; F = 0.005, P = 0.70).
In 1991, the numbers per sample on the first sampling date were eight times greater with clean cultivation (mean = 0.83 ± 0.22) than with living mulches (0.10 ± 0.05; F = 13.5, P = 0.001); by season's end the numbers were equivalent (clean cultivated mean = 0.50 ± 0.20; living mulch mean = 0.42 ± 0.15; F = 0.20, P = 0.89). Fertilizer effects were not significant in either season, with no interaction between cover crop and fertilizer factors (P>0.05). This pattern of early-season differences between treatments followed by mid- to late-season equivalencies is illustrated by catches of winged aphids in yellow water traps in 1991. On the first date, the numbers in clean-cultivated plots were 13 times greater than in living mulch plots over both fertilizer treatments (P<0.0001, fig. 2). By the second sampling date, the number of aphids trapped in the living mulches was equivalent to that counted in the clean-cultivation/compost treatment (P = 0.07); no differences were found among treatments by the third sampling date (P>0.10).
Light reflectance
The presence of cover crops between broccoli crop rows reduced early-season light reflectance at certain spectral wavebands. Compared with clean-cultivation plots, living mulch plots reflected lower levels of blue-green, yellow-orange and orange-red light in fall 1990 (fig. 3, 28 days after planting; P<0.01) and lower levels of blue-green, green, yellow and orange light in summer 1991 (fig. 4, 22 days after planting; P<0.01), with no interaction between cover crop and fertilizer factors. As the season progressed, light-reflectance patterns converged among those treatments that provided an equivalent degree of soil cover, whether that cover was from the cover crop, the increased broccoli canopy or a combination of the two. At the end of each season, broccoli in the clean-cultivated/compost treatment reflected higher-intensity light (blue-green, yellow-orange, green and orange-red in fall 1990, fig. 3, 70 days after planting; and green in summer 1991, fig. 4, 52 days after planting; P<0.01) than all other treatments. Late-season light reflectances were highest in the clean-cultivated/compost treatment either because low leaf areas prevented the broccoli canopy from fully covering the soil or because of leaf nitrogen and water stress.
Numbers of alates trapped in yellow water pans correlated positively with reflected light intensity from clean-cultivated plots in the blue-green, yellow and orange wavebands (coefficient of determination [R2] = 0.50, 0.80 and 0.56, respectively; P<0.05).
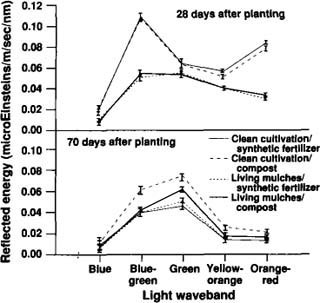
Fig. 3. Mean light energy (± MSE) reflected from broccoli plants and their backgrounds at specified wavebands in the visible light spectrum, 28 and 70 days after planting, fall 1990.
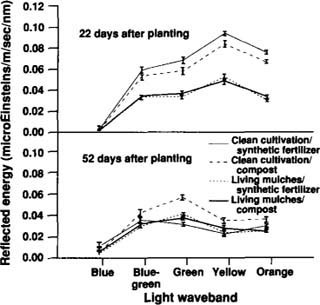
Fig. 4. Mean light energy (± MSE) reflected from broccoli plants and their backgrounds at specified wavebands in the visible light spectrum, 22 and 52 days after planting, summer 1991.
Parasitism
Season-wide parasitism in 1990 was significantly higher under clean cultivation (17.8 ± 2.4) than with living mulches (3.5 ± 2.2; F = 12.0, P = 0.003). Fertilizer had no effect, and there was no interaction between cover crop and fertilizer factors (P>0.05). In 1991, no differences were found among treatments, and parasitism over all treatments averaged 13.7%. There were no differences in aphid population growth rates among treatments in either season and no correlation between growth rates and leaf nitrate levels (P>0.05).
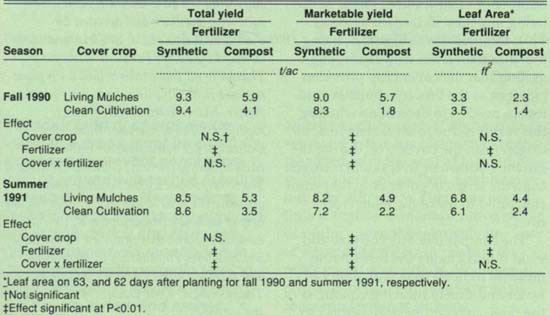
TABLE 2. Estimated gross broccoli yields and late-season, per-plant leaf area by cover crop and fertilizer level
Leaf area and yield
Neither broccoli leaf area nor yield was negatively affected by the presence of the cover crop in either season. However, in summer 1991 late-season leaf area in the living mulches was 31% higher than in the clean-cultivated control, and total yield in living mulches/compost was 53.7% higher than in clean cultivation/compost. Compared with synthetic fertilization, application of compost lowered leaf area by 79% and total yield by 87% in fall 1990, and it lowered leaf area by 62% in summer 1991. In summer 1991, marketable yield was 12% greater in living mulches/synthetic fertilizer than in clean cultivation/synthetic fertilizer. Marketable yield was 216% (fall 1990) and 122% (summer 1991) higher in living mulches/compost than in clean cultivation/compost.
Why the compost had such a deleterious effect on broccoli growth and yield is uncertain, but most likely it has to do with the lack of available nitrogen in the compost compared with the fully available nitrogen in the synthetic fertilizer. When compost is first applied, not all of the nitrogen it contains is available for plant uptake; nitrogen is released as the compost decays. The rate of release depends on temperature, soil type and the carbon-to-nitrogen ratio of the compost, among other things. Although compost was applied to the subplots three times within 1 year (summer and autumn 1990 and summer 1991), apparently there was not enough to supply levels of available nitrogen equivalent to those in the fully soluble synthetic fertilizer. The study ended before the long-term effects could be analyzed.
Conclusions
These studies show that living mulches can reduce levels of cabbage aphid infestation in harvested broccoli heads relative to broccoli grown without a cover crop. Differences in infestation levels between living mulches and clean cultivation are best explained by differences in aphid colonization rates. The correlation between light intensity and numbers of alates suggests that the lower-intensity light reflected from broccoli grown with living mulches is less attractive to incoming aphids than the higher-intensity light reflected from clean-cultivated broccoli. The strong association between numbers of alates and intensity in the yellow waveband is interesting given that the cabbage aphid is known to be attracted to the color yellow.

Tbp, broccoli /white clover living mulch, 70 days after planting, fall 1990. Plants fertilized with compost are on the right half of the plot, while those given synthetic fertilizer are on the left. Bottom, broccoli grown under clean cultivation, 70 days after planting, fall 1990. Plants fertilized with compost are on the left half of the plot, while those given synthetic fertilizer are on the right.
Plant nutrition appears to play a very important role in aphid infestation of broccoli heads. Infestation was highest under clean cultivation when compost was used as fertilizer and the plants lacked nitrogen. Although low nitrogen did not have a measurable effect on aphid population growth, aphids may have been more attracted to nitrogen-stressed flower buds. The difference between living mulches and clean cultivation in percentage of marketable (aphid-free) broccoli yield represents the true aphid control advantage of the living mulch system. Although living mulches produced higher marketable yields than clean cultivation, differences were more extreme under conditions of low nitrogen.
This is the first study in which a mechanically suppressed living mulch providing such a high level of soil coverage did not result in a smaller main crop size and yield relative to clean cultivation. The low availability of nitrogen in the compost plots did not lead to competition between the cover crop and the broccoli for soil nitrogen. Under this low-nitrogen regime, broccoli leaf area and yield in living mulches were either equal to or higher than leaf area and yield under clean cultivation, indicating that nitrogen was not a growth-limiting or yield-limiting factor in the living mulches. Indeed, in summer 1991, total yield was higher in living mulches than clean cultivation, suggesting that the cover crop provided nitrogen to the broccoli. Further studies are needed to determine whether this result is due to an exchange of nitrogen from the cover crop during the growing season or to residual nitrogen from the strips of cover crop that were cultivated to make the main crop rows.
The comparable broccoli size and yield found among the three living mulches indicate that availability of water and timely mowing equalized the three cover crops with respect to competition for water and light. Although we did not measure evapo-transpiration rates among the treatments, other researchers have found evidence that water stress reduces main crop yields in a living mulch. The tradeoff in relying on a living mulch to lower insecticide needs may be higher water use. This remains to be tested.
Moreover, we found that unless the cover crop was mowed early, severe competition for light ensued between the cover crop and the young broccoli transplants. And although it was possible to suppress the cover crop mechanically, it cost a great deal of labor. If this cropping system is to be implemented optimally, machinery must be developed to mow between the crop rows. We believe that this is feasible, given the recent development of other equipment designed for use in inter-planting systems, such as berm mowers for cover crops in orchards and vineyards.
Livina mulches suppress aphids in broccoli

