Calag Archive
Calag Archive
Imported parasitic wasp helps control red gum lerp psyllid
Publication Information
California Agriculture 59(4):229-235. https://doi.org/10.3733/ca.v059n04p229
Published October 01, 2005
PDF | Citation | Permissions
Abstract
The red gum lerp psyllid is an insect native to Australia, where it feeds upon eucalyptus species. Since 1998 this psyllid has spread throughout California, resulting in millions of dollars in damage and control costs. To help suppress the red gum lerp psyllid, a biological control program was initiated and a psyllid-specific parasitic wasp was imported from Australia in 1999 and released in 2000. In most coastal regions this biological control agent has provided substantial control, but in some interior regions the psyllid still remains a problem. Researchers are continuing their investigations to determine if full statewide suppression will be realized eventually, or if further importation of new parasitoid species is needed.
Full text
Eucalyptus trees and shrubs, valued for their ability to flourish in arid regions and their varied horticultural uses, have been a familiar feature of California's urban and rural landscapes since they were first introduced from their native Australia more than 150 years ago. Until recently, eucalyptus trees in California were relatively free from damaging insect pests. Most of California's native insects cannot feed on eucalyptus, which is well protected from herbivores by chemicals such as distasteful essential oils (which are familiar to anyone who has smelled the strong odor of the leaves). The Australian insects that have adapted to feed on eucalyptus were not transported to California with earlier shipments of plant propagation material, usually in the form of seeds. This began to change in the early 1980s and at least 15 eucalyptus-feeding insect species from Australia were accidentally introduced and are now established in California (Paine and Millar 2002). While eucalyptus trees may be unwanted in some areas because they crowd out native vegetation, their extensive value in many other locations led to efforts to control the psyllid.
River red gum (Eucalyptus camal-dulensis) is among the most commonly planted shade and windbreak trees in California and is also grown commercially for fuel wood and fiber (Cockerham 2004). The red gum lerp psyllid (Glycaspsis brimblecombei Moore; Hemiptera: Psylloidea) was first found on river red gum in June 1998 in Los Angeles County and had spread throughout the state by 2000, and throughout Mexico and parts of Florida by 2002. In Australia there are a number of eucalyptus species that the red gum lerp psyllid can feed on, but in California the only favored eucalyptus species present is the river red gum; the forest red gum (E. tereticornis) and flooded gum (E. rudis), both also in California, are less-favored trees that the psyllid can feed on as well.

The parasitoid Psyllaphaegus bliteus has been released throughout California to control the red gum lerp psyllid, a pest of eucalyptus. Above, an adult P. bliteus uses its ovipositor to place an egg inside the red gum lerp psyllid nymph. The parasitoid develops inside the psyllid nymph, which typically does not show any signs of parasitism until the nymph reaches the fifth instar, when the parasitoid pupa - far left, white body and left, dark body - can be seen through the mummified psyllid.
Red gum lerp psyllid nymphs build white conical shelters called lerps from excreted honeydew and waxes, and live underneath these structures. The nymphs feed by sucking plant sap from leaves. The accumulation of the sticky lerps and honeydew on leaves and under infested trees creates a nuisance, while heavy infestations lead to defoliation, branch dieback and occasionally tree death (Paine et al. 2000).
The first attempts to control red gum lerp psyllid focused on the use of systemic insecticides, mainly to target heavy infestations on particularly valuable trees. The proper timing of treatments was difficult to determine and control was not always achieved (Paine et al. 2000). The obvious impracticality of using insecticide treatments on trees throughout the state led us to investigate more sustainable options. We first investigated whether any predators already present in California could provide control. Lady beetles (Hippodamia), green lacewings (Chrysoperla), minute pirate bugs (Orius) and syrphid flies feed on adult and immature red gum lerp psyllids (Erbilgin et al. 2004). However, even when present in large numbers, these predators did not provide adequate control.

Clockwise from top left: Karen Sime, UC Berkeley postdoctorate researcher, checks a caged eucalyptus leaf for evidence of P. bliteus activity and parasitism rates; the late Don Dahisten (first author) was a leading UC researcher (1966–2003) in biological control of urban and forest pests (shown in 1974); small discs coated with a light oil were used to capture and sample populations of adult red gum lerp psyllid and P. bliteus red gum lerp psyllids feed on eucalyptus leaves, building up to such high densities that the accumulation of psyllids and honeydew causes sooty molds, defoliation and even tree death; in the San Joaquin Valley (Tulare County), a dead red gum eucalyptus (left) near an undamaged and uninfested blue gum eucalyptus (right) demonstrates the psyllid's feeding preferences.

Life stages of the red gum lerp psyllid include, (left to right) large nymph, row of eggs, winged adult and small lerp (the protective covering produced by nymphs).
In Australia, red gum lerp psyllid populations are held in check in large part by species of parasitic wasps that specifically attack them and their close relatives. Other parasitoid species imported from Australia had successfully controlled earlier outbreaks of other introduced Australian psyllid species in California, including the blue gum psyllid (Dahlsten et al. 1998) and the eugenia psyllid (Dahlsten et al. 1995). Classical biological control appeared, therefore, to be the most promising approach for controlling the red gum lerp psyllid. We report here on a large collaborative effort between UC, the California Department of Food and Agriculture (CDFA) and research scientists in Australia.
Prerelease psyllid sampling
Before the biological control program began, we gathered detailed information about red gum lerp psyllid populations throughout California. Beginning in July 1999, sample sites were established in Alameda, Santa Clara, Monterey, Los Angeles and San Diego counties. By July 2002, we had established 32 sample sites, with at least one site located in every California county in which red gum lerp psyllid had been reported.
Sampling psyllid populations accurately can be difficult. The most accurate way of measuring densities and damage levels is to count psyllid nymphs on leaves. This method, however, is time-consuming and impractical for the large number of sample sites and frequent sampling dates needed to follow psyllid and natural-enemy population dynamics throughout the state. We therefore investigated whether red gum lerp psyllid populations could be tracked with sticky traps, which had been used for the blue gum and eugenia psyllid programs (Dahlsten et al. 1998). The traps consisted of transparent 4-inch (10-centimeter) plastic disks coated with a thin layer of motor oil additive and clipped over a yellow backing. At each site, 10 to 12 traps were hung in eucalyptus trees and changed weekly.
In 1999 and 2000, we tested trap accuracy at two sites, one in Northern California (Alameda County) and the other in Southern California (Los Angeles County). Near each trap at these sites, two 12-inch (30-centimeter) foliage samples were collected every 3 weeks (20 to 24 samples per site per sample date). We found a good correlation between the mean number of adult female psyllids per sticky trap and the mean number of psyllid eggs per leaf sample (P < 0.01, r2 = 0.82; Paine et al. [2000]), indicating that the sticky-trap counts provided a good estimate of psyllid activity in eucalyptus. Thereafter, we relied exclusively on the sticky-trap technique to monitor the psyllids. In addition, we used the same traps to monitor parasitoid populations after we began releasing them.

Fig 1. P. bliteus oviposition success in different host developmental stages, as indicated by the percentage egg deposition (± SEM) of parasitized red gum lerp psyllids, was significantly different (F = 12.48, df = 4,25, P < 0.001). Above each bar, means followed by different letters are significantly different (Tukey's HSD test, P < 0.05). Source: Daane et al. (2005).
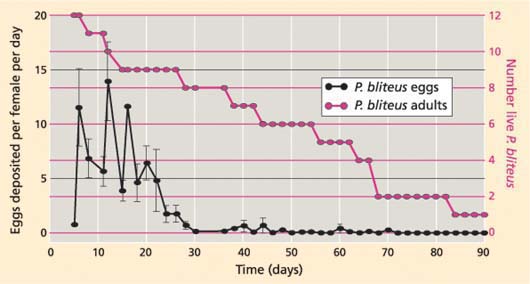
Fig 2. P. bliteus lifetime fecundity under glasshouse conditions, as estimated by egg deposition with an overabundant host supply. Source: Daane et al. (2005).
Initially, we counted both male and female adult psyllids per trap by sample week. However, we noticed strong seasonal changes in the relative proportions of adult females and males. For example, female psyllids typically had the highest populations in the summer months and dropped significantly the rest of the year. Because their numbers are most closely associated with egg-laying (and thus nymphal activity and damage), we plotted only the adult female psyllids caught on the traps. At each monitoring site, we reported the averages of 10 to 12 traps.
Finding, importing parasitoids
To find promising parasitoids, mummified red gum lerp psyllids were field-collected and shipped from southern Australia to the UC Berkeley Quarantine Facility in August 1999 (the “mummified” psyllid is visibly dead, killed by the internal parasitoid that is nearing completion of its development). This region, near Adelaide, has a Mediterranean climate, with temperatures similar to California's coastal regions. From the field-collected psyllids, we reared eight species of parasitoids in the genus Psyllaephagus (Hymenoptera: Encyrtidae). Of these, two were hyperparasitoids (which attacked the beneficial “primary” parasitoids) that attacked the other Psyllaephagus species, and most others failed to propagate in the insectary. The remaining species (Psyllaephagus bliteus Riek) did well in culture and was selected for release after experiments showed that it specifically attacked the red gum lerp psyllid when tested against three other psyllid species (Eugenia, blue gum and mela-leuca) (Paine et al. 2000).
Parasitoid biology
To facilitate insectary rearing and field release, we collected basic biological information on P. bliteus (Daane et al. 2005). First, we determined which stages of the host (red gum lerp psyllid) were preferred by the female parasitoids (P. bliteus) for oviposition. Potted eucalyptus trees were infested with 300 to 500 psyllid nymphs, with the population composed of all five nymphal stages in similar proportions, and isolated in organdy sleeve cages with 15 to 20 adult female P. bliteus. After 24 hours, all psyllids were collected and cleared in chloralphenol, which makes any P. bliteus eggs inside the psyllid body visible under a dissecting microscope. The number of eggs and the psyllid stages were recorded. We also used similar methods to investigate P. bliteus larval development.
We collected the psyllids every 3 to 4 days after exposure to female wasps, cleared them in chloralphenol, and then recorded both the presence of P. bliteus eggs or larvae and the developmental stages of parasitized psyllids. There were five host preference and four larval development replicates; the treatment means were separated using Tukey's HSD test. Our results showed that P. bliteus can oviposit into psyllid nymphs of any age, but that they usually parasitize third and fourth instars (fig. 1). In our studies, regardless of the stage of the host exposed to P. bliteus for oviposition, the parasite larvae did not fully develop until after the host reached the fifth (last) instar. In addition, adult female wasps also occasionally killed psyllid nymphs by host feeding, stabbing them with their ovipositors and drinking the body fluids leaking out from under the lerp. Usually younger nymphs are attacked but we observed this host feeding on all stages.
Longevity and fecundity
Adult P. bliteus longevity and fecundity were also determined. Newly emerged and mated female P. bliteus were individually isolated in clear plastic tubes that each enclosed a single infested leaf on a potted river red gum tree in the glasshouse (71.6 ± 3°F). Each leaf was infested with 10 to 30 psyllids, mostly third instars. Each female P. bliteus was transferred to a new leaf every 2 days throughout her lifetime. After each transfer, the parasitoid-exposed psyllids were cleared in chloralphenol, and the presence of P. bliteus eggs was recorded. Under these conditions, we found that average female P. bliteus longevity was 60.4 ± 6.4 days and average lifetime egg deposition was 125.7 ± 24.6 eggs per female (range 34 to 302). Most eggs (88.1%) were deposited during the initial 22 days, although one parasitoid deposited eggs up to 70 days after emergence (fig. 2).
These results have implications for insectary operations and release strategies in classical biological control programs. Although adults may survive for long periods, most egg deposition occurs early in the adult's lifetime. Insectary colonies should therefore be supplied with the needed number of third- or fourth-instar psyllids for an oviposition period of 2 to 3 weeks. In addition, first or second instars should also be provided because they are used for host feeding. Finally, the parasitic wasps can be released to the field shortly after emergence and mating, as high rates of egg deposition begin immediately.

TABLE 1. P. bliteus releases by California county, 2000 to 2003, show the widespread collaborative effort to release and establish the imported red gum lerp psyllid parasitoid
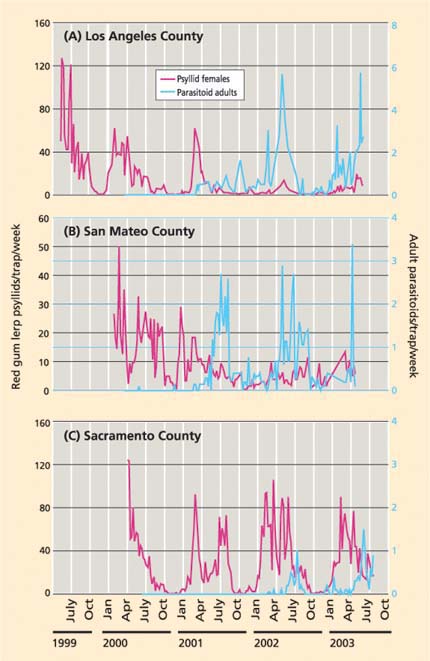
Fig 3. Red gum lerp psyllid adult females and P. bliteus parasitoid adults (both mean per trap per week) and parasitoid release dates in a single site each in (A) eastern Los Angeles County, (B) San Mateo County (parasitoids originally released in nearby counties) and (C) southern Sacramento County.
Parasitoid release and impact
For field release, most parasitoids were reared at the CDFA Biological Control Facility (Sacramento), with smaller numbers reared at the UC Berkeley Insectary and Quarantine Facility. Release of P. bliteus began in June 2000 in Los Angeles and Alameda counties. Between September 2000 and January 2003, we released a total of 48,582 adults in 78 release sites located in 42 counties throughout the state (1,156 ± 154 per county, range 50 to 4,016) (table 1).
Parasitoids were recovered in sticky traps as early as 8 weeks after initial release. Recovery in traps occurred earliest in the Central Coast sites, followed by the Southern California, North Coast and Central Valley sites (table 2). Psyllid densities typically peaked between August and October, and these peak periods were used to compare annual changes at each site. Eastern Los Angeles County is an example of a site with a marked decrease in psyllids and an increase in parasitoids (fig. 3A). Peak psyllid counts dropped from more than 100 in 1999 (before the wasp release) to fewer than 20 females per trap per week in 2003 (after the second wasp release). During the same period, trap catches of P. bliteus increased for 3 years after the initial release in June 2000 and then showed steady seasonal cycles. A similar pattern of psyllid decrease and parasitoid increase was found farther north, in San Mateo County (parasitoids were originally released in nearby counties) (fig. 3B). In contrast, P. bliteus has to date had less effect on psyllid densities in some interior sites. For example, 3 years after parasitoid releases at one Sacramento County location, parasitoid levels were relatively low and psyllid numbers remained high (fig. 3C).
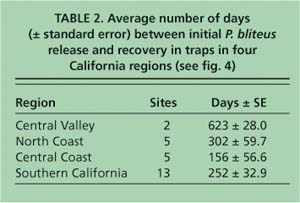
TABLE 2. Average number of days (± standard error) between initial R. bliteus release and recovery in traps in four California regions (see fig. 4)
To summarize the changes in psyllid density across all 32 monitored sites, we compared the average peak densities of psyllids per trap in 2003 to those in years before the parasitoid was established (1999 to 2001). The results of this comparison, grouped by region, indicate a postrelease drop in peak psyllid densities of 78.6%, 59.5% and 44.8% in the southern, central and northern coastal regions, respectively. There was no change in peak psyllid densities in the Central Valley sites. The postrelease rate of increased parasitoid activity was estimated by calculating the average time from the release to a 50% or more decrease in peak psyllid density on the monitoring traps. At most coastal sites (18 out of 23) the average time to a 50% decrease in psyllid density was 13.2 ± 1.2 months after the initial parasitoid release. By comparison, at the nine Central Valley sites monitored, there was little or no detectable effect of P. bliteus on the psyllid population densities during the 2001 to 2003 sampling period.
Coastal versus interior sites
The differences in parasitoid effectiveness between coastal and interior sites were most likely due to the great climatic differences between the two regions. The coastal region has relatively mild summer and winter temperatures, while the interior valleys have much cooler winters and warmer summers. To date, the Central Valley sites lag behind the coastal sites in two measures of parasitoid impact: the time to establishment of parasitoid populations and the overall impact of the parasitoids on psyllid populations. To account for these discrepancies, we compared the performance of the parasitoids at coastal and interior sites that were paired by latitude using three measures: (1) parasitism rates, (2) the ability of a single wasp to parasitize psyllids and (3) the longevity of adult female wasps outdoors.
In the first study, three pairs of coastal and interior monitoring sites were sampled in July 2003: (1) Sonoma and Sacramento counties, (2) Alameda and Fresno counties and (3) San Luis Obispo and Bakersfield counties. Two 11.8-inch (30-centimeter) branch tips were taken from each site and up to 50 nymphs from each sample were dissected to check for parasitism. We found that the average parasitism rate at coastal sites (Sonoma, Alameda and San Luis Obispo counties) (29.7% ± 6.6%) was significantly higher than the average parasitism rate (1.2 ± 0.9%) at the interior sites (Sacramento, Fresno and Kern counties) (t-test, P < 0.05, n = 8).
In a second study, three to four mesh sleeve cages were fixed to river red gums at two paired sites (Alameda and Sacramento counties) in July 2003. These sites represented the coastal and interior temperature regimes at the same latitude; mean average and high temperatures for the cities of Berkeley (Alameda County) and Sacramento (Sacramento County), where the study was conducted, are 63°F and 71°F, and 74°F and 91°F, respectively. Approximately 100 psyllid eggs were placed in each cage, and the resulting nymphs then settled on leaves and began feeding. When the psyllids had reached the third instar, a single female wasp was released into each cage. The cages were removed from the field 2 weeks later and all psyllids inside were dissected to check for parasitism. Average parasitism rates in the cages were far higher at the Berkeley site (34.1% ± 9.31%) than at the Sacramento site (1.0% ± 1.0%) (2-tailed t-test, df = 14, t = 2.145, P = 0.003).
In a third temperature study, we compared the longevity of adult female parasitoids held in glass vials at the sites in Berkeley and Sacramento during summer 2004. During a period in June when temperatures were cool in Berkeley but warm in Sacramento, the wasps lived significantly longer in Berkeley (14.9 ± 2.4 days versus 5.8 ± 0.7 days) (2-tailed t-test, df = 3, unpaired t = 3.182, P = 0.037). During the July and September test periods, temperatures were similar at the two sites (with unusually cool weather in Sacramento in that part of July) and the wasps' longevity was similar at both sites (averaging 12 to 15 days).
The wasps performed poorly in the Central Valley compared to coastal areas. Parasitism rates were lower overall in the field, and individual wasps laid fewer eggs. A possible explanation for these differences is the shorter lifespan of the wasps in the summer heat. In our experiments on the basic biology of the parasitoids, we found that they laid eggs for several weeks in the greenhouse, which has mild, cool conditions similar to the ambient conditions in Berkeley in the summer. However, in warmer conditions, the wasps did not live as long and therefore laid fewer eggs over their lifetimes. The relatively poor performance of the wasps in the summer in the Central Valley, when psyllid numbers build up to their seasonal peaks, helps explain why we have observed longer times for wasp establishment in the interior and, to date, less impact on psyllid populations.
Parasitoid impact
By 2003, P. bliteus had been recovered at 29 of the 32 sites monitored throughout the season with sticky traps. Field-produced parasitoids far outnumbered insectary production and, for this reason, managed releases were discontinued. To provide a geographically comprehensive summary of parasitoid establishment, we surveyed 55 former release sites throughout California from mid-August through October 2003 (Roltsch et al. 2004). At each site, 15 branch terminals (11.8 to 15.7 inches long) were randomly collected from three or more trees. On each branch, 30 leaves were randomly selected and the numbers of exit holes (round holes that the adult parasitoid chews in the lerp when it exits), as well as healthy and visibly parasitized psyllids, were recorded by life stage.
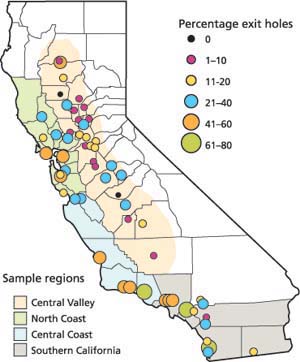
Fig 4. Percentage of large lerps with parasitoid emergence exit holes during fall survey, 2003 (Roltsch et al. 2004). California county borders demark the four regional sampling zones (except for the Central Valley).
Parasitoids were recovered at all but two of 55 locations (fig. 4). At several locations, levels of parasitoid activity, as indicated by the proportion of large lerps containing exit holes, were strikingly higher than those found in 2002, several months after the initial parasitoid releases, using similar survey techniques. For example, while P. bliteus was extremely rare at the Solano County site in 2002 (after two releases prior to this sampling), in October 2003 the parasitoid was common there. On average over all sites sampled, there were parasitoid exit holes in more than one-fifth (22%) of the large lerps. We note that while exit holes in the lerps provide an indication of parasitoid activity, this may not be as accurate as dissecting live psyllids to determine the percentage that are parasitized. Still, the survey confirms that the released P. bliteus have established in almost all regions and that parasitoid activity appears to be increasing annually.
Future biocontrol programs
The red gum lerp psyllid now appears to be under control in most coastal regions of California due to the introduction of P. bliteus. Suppression is best at coastal sites and lower in some parts of the Central Valley. Our field and laboratory studies suggest that the hot summer temperatures found in the interior regions may reduce parasitoid impact. Still, as psyllid numbers have dropped, the defoliation and death of eucalyptus trees due to the psyllid have been reduced. Of key importance for future control efforts is the observation that P. bliteus appears to be well established throughout California, including the interior locations. Therefore, although current P. bliteus densities and parasitism rates are low in the interior, their impact may continue to increase there, albeit more slowly than in coastal regions. We will continue to monitor the red gum lerp psyllid and the parasitoid populations to determine if it will be necessary to import either heat-adapted P. bliteus populations or additional Psyllaephagus species to improve biological control in California's interior.
Imported parasitic wasp helps control red gum lerp psyllid

